Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body (24 page)
Read Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body Online
Authors: Neil Shubin

GENES

Arendt’s discovery leads to yet another question. It is one thing for eyes to share common parts, but how can eyes that look so different—such as those of worms, flies, and mice—be closely related? For the answer, let us consider the genetic recipe that builds eyes.
At the turn of the twentieth century, Mildred Hoge was recording mutations in fruit flies when she found a fly that had no eyes whatsoever. This mutant was not an isolated case, and Hoge discovered that she could breed a whole line of such flies, which she named
eyeless.
Later, a similar mutation was discovered in mice. Some individuals had small eyes; others lacked whole portions of the head and face, including their eyes. A similar condition in humans is known as aniridia; affected individuals are missing large pieces of their eyes. In these very different creatures—flies, mice, and humans—geneticists were finding similar kinds of mutants.
A breakthrough came in the early 1990s, when laboratories applied new molecular techniques to understand how eyeless mutants affected eye development. Mapping the genes, they were able to localize the bits of DNA responsible for the mutations. When the DNA was sequenced, it turned out that the fly, mouse, and human genes responsible for eyelessness had similar DNA structures and sequences. In a very real sense, they are the same gene.
What did we learn from this? Scientists had identified a single gene that, when mutated, produced creatures with small eyes or no eyes at all. This meant that the normal version of the gene was a major trigger for the formation of eyes. Now came the chance to do experiments to ask a whole other kind of question. What happens when we mess with the gene, turning it on and off in the wrong places?
Flies were an ideal subject for this work. During the 1980s, a number of very powerful genetic tools were developed through work on flies. If you knew a gene, or a DNA sequence, you could make a fly lacking the gene or, the reverse, a fly with the gene active in the wrong places.
Using these tools, Walter Gehring started playing around with the
eyeless
gene. Gehring’s team was able to make the
eyeless
DNA active pretty much anywhere they wanted: in the antenna, on the legs, on the wings. When his team did this, they found something stunning. If they turned on the
eyeless
gene in the antenna, an eye grew there. If they turned on the
eyeless
gene on a body segment, an eye developed there. Everywhere they turned on the gene, they would get a new eye. To top it all off, some of the misplaced eyes showed a nascent ability to respond to light. Gehring had uncovered a major trigger in the formation of eyes.
Gehring didn’t stop there; he began swapping genes between species. They took the mouse equivalent of
eyeless, Pax 6,
and turned it on in a fly. The mouse gene produced a new eye. And not just any eye—a fly eye. Gehring’s lab found they could use the
mouse
gene to trigger the formation of an extra
fly
eye anywhere: on the back, on a wing, near the mouth. What Gehring had found was a master switch for eye development that was virtually the same in a mouse and a fly. This gene,
Pax 6,
initiated a complex chain reaction of gene activity that ultimately led to a new fly eye.
We now know that
eyeless,
or
Pax 6,
controls development in everything that has eyes. The eyes may look different—some with a lens, some without; some compound, some simple—but the genetic switches that make them are the same.
When you look into eyes, forget about romance, creation, and the windows into the soul. With their molecules, genes, and tissues derived from microbes, jellyfish, worms, and flies, you see an entire menagerie.
CHAPTER TEN
EARS
T
he first time you see the inside of the ear is a letdown: the real machinery is hidden deep inside the skull, encased in a wall of bone. Once you open the skull and remove the brain, you need to chip with a chisel to remove that wall. If you are really good, or very lucky, you’ll make the right stroke and see it—the inner ear. It resembles the kind of tiny coiled snail shell you find in the dirt in your lawn.
The ear may not look like much, but it is a wonderful Rube Goldberg contraption. When we hear, sound waves are funneled into the outside flap, the external ear. The sound waves enter the ear and make the eardrum rattle. The eardrum is attached to three little bones, which shake along with it. One of these ear bones is attached to the snail-shell structure by a kind of plunger. The shaking of the ear bone causes the plunger to go up and down. This causes some gel inside the snail shell to move around. Swishing gel bends nerves, which send a signal to the brain, which interprets it as sound. Next time you are at a concert, just imagine all the stuff flying around in your head.
This structure allows us to distinguish three parts to the ear: external, middle, and inner. The external ear is the visible part. The middle contains the little ear bones. Finally, the inner ear consists of the nerves, the gel, and the tissues that surround them. These three components of ears enable us to structure our discussion in a very convenient way.
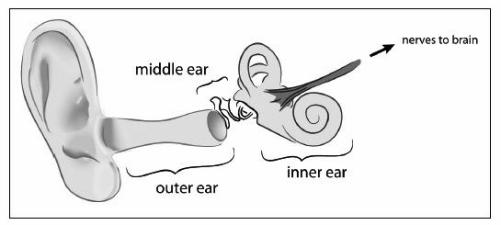
Of the three parts of our ear—the outer, middle, and inner—the inner ear is the most ancient and the part that controls the nerve impulses sent to the brain.
The part of the ear we can see, the flap on which we hang our glasses, is a relatively new evolutionary addition to bodies. Confirm this on your next trip to the aquarium or zoo. How many sharks, bony fish, amphibians, and reptiles have external ears? The pinna—the flap of the external ear—is found only in mammals. Some amphibians and reptiles have visible external ears, but they have no pinna. Often the external ear is only a membrane that looks like the top of a drum.
The elegance of our connection to sharks and bony fish is revealed when we look inside our ears. Ears might seem an unlikely place for a human-shark connection, especially since sharks don’t have ears. But the connection is there. Let’s start with the ear bones.
THE MIDDLE EAR—THE THREE EAR BONES
Mammals are very special. With hair and milk-producing glands, we can easily be distinguished from other creatures. It surprises most people to learn that some of the most distinctive traits of mammals lie inside the ear. The bones of the mammalian middle ear are like those of no other animal: mammals have three bones, whereas reptiles and amphibians have only one. Fish have none at all. Where did our middle ear bones come from?
Some anatomy: recall that our three middle ear bones are known as the malleus, incus, and stapes. As we’ve seen, each of these ear bones is derived from the gill arches: the stapes from the first arch, and the malleus and incus from the second arch. It is here that our story begins.
In 1837, the German anatomist Karl Reichert was looking at embryos of mammals and reptiles to understand how the skull forms. He followed the gill arches of different species to understand where they ended up in the various skulls. As he did this again and again, he found something that appeared not to make any sense: two of the ear bones in the mammals corresponded to pieces of the jaw in the reptiles. Reichert could not believe his eyes, and his monograph reveals his excitement. As he describes the ear-jaw comparison, his prose departs from the normally staid description of nineteenth-century anatomy to express shock, even wonderment, at this discovery. The conclusion was inescapable: the same gill arch that formed part of the jaw of a reptile formed ear bones in mammals. Reichert proposed a notion that even he could barely believe—that parts of the ears of mammals are the same thing as the jaws of reptiles. Things get more difficult when we realize that Reichert proposed this several decades before Darwin propounded his notion of a family tree for life. What does it mean to call structures in two different species “the same” without a notion of evolution?
Much later, in 1910 and 1912, the German anatomist Ernst Gaupp picked up on Reichert’s work and published an exhaustive study on the embryology of mammalian ears. Gaupp provided more detail and, given the times, interpreted Reichert’s work in an evolutionary framework. Gaupp’s story went like this: the three middle ear bones reveal the tie between reptiles and mammals. The single bone in the reptilian middle ear is the same as the stapes of mammals; both are second-arch derivatives. The explosive bit of information, though, was that the two other middle ear bones of mammals—the malleus and the incus—evolved from bones set in the back of the reptilian jaw. If this was indeed the case, then the fossil record should show bones shifting from the jaw to the ear during the origin of mammals. The problem was that Gaupp worked only on living creatures and didn’t fully appreciate the role that fossils could play in his theory.
Beginning in the 1840s a number of new kinds of fossil creatures were becoming known from discoveries in South Africa and Russia. Often abundantly preserved, whole skeletons of dog-size animals were being unearthed. As they were discovered, many of them were crated and shipped to Richard Owen in London for identification and analysis. Owen was struck that these creatures had a mélange of features. Parts of their skeleton looked reptile-like. Other parts, notably their teeth, looked like mammals. And these were not isolated finds. It turns out that these “mammal-like reptiles” were the most common skeletons being uncovered at many fossil sites. Not only were they very common, there were many kinds. In the years after Owen, these mammal-like reptiles became known from other parts of the world and from several different time periods in earth history. They formed a beautiful transitional series in the fossil record between reptile and mammal.