Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body (23 page)
Read Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body Online
Authors: Neil Shubin

VISION
O
nly once in my entire career have I found the eye of a fossil creature. I wasn’t in the field on an expedition, I was in the back room of a mineral shop in a small town in northeast China. My colleague Gao Keqin and I were studying the earliest known salamanders, beautiful fossils collected from Chinese rocks about 160 million years old. We had just finished a collecting trip to some sites Gao knew about. The locations were secret, because these salamander fossils have serious monetary value for the farmers who typically find them. What makes them special is that impressions of the soft tissue, such as gills, guts, and the notochord, are often preserved. Private collectors love them because fossils of this quality are exceedingly rare. By the time we ended up at the mineral shop, Gao and I had already collected a number of really beautiful ancient salamanders of our own from his sites.
This particular mineral dealer had gotten his hands on one of the best salamander fossils of all time. Gao wanted us to see it and spent the better part of a day trying to work the deal. The whole visit had a terrifically illicit feel. Gao spent several hours smoking cigarettes with the gentleman, speaking and gesturing in Chinese. Clearly there was some bartering going on, but not knowing Chinese I had no idea what offers were being put on the table. After lots of headshaking and ultimately a big handshake, I was permitted to go to the back room and look at a fossil on the dealer’s desk. It was a stunning sight: the body of a larval salamander, no more than three inches long. In it, I could see impressions of the whole animal, all the way down to the little shells it ate as its last meal. And, for the first and only time in my career, I was staring at the eye of an ancient fossil animal.
Eyes rarely make it into the fossil record. As we’ve seen, the best candidates for preservation as fossils are the hard parts of the animals—bones, teeth, and scales. If we want to understand the history of eyes, then we can use an important fact to our advantage. There is a remarkable diversity of organs and tissues that animals use to capture light, from simple photoreceptor organs in invertebrate animals to the compound eyes of various insects and our own camera-type eye. How do we put this variation to use in understanding how our ability to see developed over time?
The history of our eyes is a lot like that of a car. Take a Chevy Corvette, for example. We can trace the history of the model as a whole—the Corvette—and the history of each of its parts. The ’Vette has a history, beginning with its origins in 1953 and continuing through the different model designs each year. The tires used on the ’Vette also have a history, as does the rubber used in making them. This supplies a great analogy for bodies and organs. Our eyes have a history as organs, but so do eyes’ constituent parts, the cells and tissues, and so do the genes that make those parts. Once we identify these multiple layers of history in our organs, we understand that we are simply a mosaic of bits and pieces found in virtually everything else on the planet.
Much of the processing of the images we see actually happens inside our brains: the role of the eye is to capture light in a way that it can be carried to the brain for processing as an image. Our eyes, like those of every creature with a skull and backbone, are like little cameras. After light from the outside enters the eye, it is focused on a screen at the back of the eyeball. Light travels through several layers as it traverses this path. First it passes through the cornea, a thin layer of clear tissue that covers the lens. The amount of light that enters the eye is controlled by a diaphragm, called the iris, which dilates and contracts by the action of involuntary muscles. The light then passes through the lens, which, as a camera does, focuses the image. Tiny muscles surround the lens; as these muscles contract, they change the lens’s shape, thus focusing images from near and far. A healthy lens is clear and made up of special proteins that give it its distinctive shape and light-gathering properties. These proteins, known as lens crystallins, are exceptionally long-lived, allowing the lens to continue functioning as we age. The screen on which all of the light is projected, the retina, is loaded with blood vessels and light receptors. These light receptors send signals to our brain that we then interpret as images. The retina absorbs the light via sensitive light-gathering cells. There are two types of such cells: one is very sensitive to light, the other less so. The more sensitive cells record only in black and white; the less sensitive cells record in color. If we look around the animal world, we can assess whether animals are specialized for daylight or night by looking at the percentages of each type of light-sensing cell in their eyes. In humans these cells make up about 70 percent of all the sensory cells in our body. That is a clear statement about how important vision is to us.
Our camera-like eye is common to every creature with a skull, from fish to mammals. In other groups of animals we find different eyes, ranging from simple patches of cells specialized to detect light, to eyes with compound lenses such as those found in flies, to primordial versions of our own eye. The key to understanding the history of our eyes is to understand the relationship between the structures that make our camera-eye and those that make all the other kinds of eyes. To do this, we will study the molecules that gather light, the tissues we use to see, and the genes that make the whole thing.
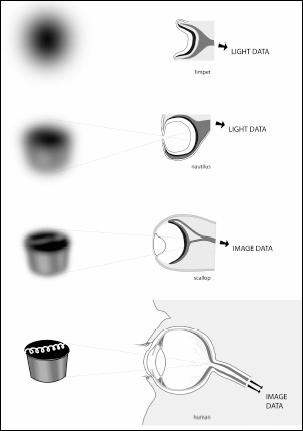
Eyes come into focus: from primitive light-capturing devices in invertebrates to our camera-type eye with a lens. As eyes evolve, visual acuity increases.
LIGHT-GATHERING MOLECULES

The really important work in the light-gathering cells happens inside the molecule that actually collects light. When this molecule absorbs light, it changes shape and breaks up into two parts. One part is derived from vitamin A, the other from a protein known as an opsin. When the opsin breaks off, it initiates a chain reaction that leads to a neuron sending an impulse to our brain. We use different opsins to see in black and white and in color. Just as an inkjet printer needs three or four inks to print in color, we need three light-gathering molecules to see in color. For black-and-white vision, we use only one.
These light-gathering molecules change shape in the light, then recharge in the dark and go back to their normal state. The process takes a few minutes. We all know this from personal experience: go from a bright place into a dark room and it is virtually impossible to see faint objects. The reason is that the light-gathering molecules need time to recharge. After a few minutes, vision in the dark returns.
Despite the stunning variety of photoreceptor organs, every animal uses the same kind of light-capturing molecule to do this job. Insects, humans, clams, and scallops all use opsins. Not only can we trace the history of eyes through differences in the structure of their opsins, but we have good evidence that we can thank bacteria for these molecules in the first place.
Essentially, an opsin is a kind of molecule that conveys information from the outside of a cell to the inside. To pull off this feat, it needs to carry a chemical across the membrane that encircles a cell. Opsins use a specialized kind of conductor that takes a series of bends and loops as it travels from the outside to the inside of the cell. But this twisted path the receptor takes through the cell membrane is not random—it has a characteristic signature. Where else is this twisted path seen? It is identical to parts of certain molecules in bacteria. The very precise molecular similarities in this molecule suggest a very ancient property of all animals extending all the way to our shared history with bacteria. In a sense, modified bits of ancient bacteria lie inside our retinas, helping us to see.
We can even trace some major events in the history of our eyes by examining opsins in different animals. Take one of the major events in our primate past, the development of rich color vision. Recall that humans and our closest ape relatives, the Old World monkeys, have a very detailed kind of color vision that relies on three different kinds of light receptors. Each of these receptors is tuned to a different kind of light. Most other mammals have only two kinds of receptors and so cannot discriminate as many colors as we can. It turns out that we can trace the origin of our color vision by looking at the genes that make the receptors. The two kinds of receptors most mammals have are made by two kinds of genes. Of our three receptor-making genes, two are remarkably like one of those in other mammals. This seems to imply that our color vision began when one of the genes in other mammals duplicated and the copies specialized over time for different light sources. As you’ll remember, a similar thing happened with odor receptor genes.
This shift may be related to changes in the flora of the earth millions of years ago. It helps to think what color vision was likely good for when it first appeared. Monkeys that live in trees would benefit because color vision enabled them to discriminate better among many kinds of fruits and leaves and select the most nutritious among them. From studying the other primates that have color vision, we can estimate that our kind of color vision arose about 55 million years ago. At this time we find fossil evidence of changes in the composition of ancient forests. Before this time, the forests were rich in figs and palms, which are tasty but all of the same general color. Later forests had more of a diversity of plants, likely with different colors. It seems a good bet that the switch to color vision correlates with a switch from a monochromatic forest to one with a richer palette of colors in food.
TISSUES
Animal eyes come in two flavors; one is seen in invertebrates, the other in vertebrates, such as fish and humans. The central idea is that there are two different ways of increasing the light-gathering surface area in eye tissue. Invertebrates, such as flies and worms, accomplish this by having numerous folds in the tissue, while our lineage expands the surface area by having lots of little projections extending from the tissue like tiny bristles. A host of other differences also relate to these different kinds of designs. Lacking fossils at the relevant phase of history, it would seem that we would never be able to bridge the differences between our eyes and those of invertebrates. That is, until 2001, when Detlev Arendt thought to study the eyes of a very primitive little worm.
Polychaetes are among the most primitive living worms known. They have a very simple segmented body plan, and they also have two kinds of light-sensing organs: an eye and, buried under their skin, a part of their nervous system that is specialized to pick up light. Arendt took these worms apart both physically and genetically. Knowledge of the gene sequence of our opsin genes and the structure of our light-gathering neurons gave Arendt the tools to study how polychaetes are made. He found that they had elements of both kinds of animal photoreceptors. The normal “eye” was made up of neurons and opsins like the eye of any invertebrate. The tiny photoreceptors under the skin were another matter altogether. They had “vertebrate” opsins and cellular structure even with the little bristle-like projections, but in primitive form. Arendt had found a living bridge, an animal with both kinds of eyes, one of which—our kind—existed in a very primitive form. When we look to primitive invertebrates, we find that the different kinds of animal eyes share common parts.