Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body (25 page)
Read Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body Online
Authors: Neil Shubin

Until 1913, embryologists and paleontologists were working in isolation from one another. At this time, the American paleontologist W. K. Gregory, of the American Museum of Natural History, saw an important link between Gaupp’s embryos and the African fossils. The most reptilian of the mammal-like reptiles had only a single bone in its middle ear; like other reptiles, it had a jaw composed of many bones. Something remarkable was revealed as Gregory looked at the successively more mammalian mammal-like reptiles, something that would have floored Reichert had he been alive: a continuum of forms showing beyond doubt that over time the bones at the back of the reptilian jaw got smaller and smaller, until they ultimately lay in the middle ear of mammals. The malleus and incus did indeed evolve from jawbones. What Reichert and Gaupp observed in embryos was buried in the fossil record all along, just waiting to be discovered.
Why would mammals need a three-boned middle ear? This little linkage forms a lever system that allows mammals to hear higher-frequency sounds than animals with a single middle ear bone. The origin of mammals involved not only new patterns of chewing, as we saw in Chapter 4, but new ways of hearing. In fact, this shift was accomplished not by evolving new bones per se, but by repurposing existing ones. Bones originally used by reptiles to chew evolved in mammals to assist in hearing.
So much for the malleus and incus. Where, though, does the stapes come from?
If I simply showed you an adult human and a shark, you would never guess that this tiny bone deep inside a human’s ear is the same thing as a large rod in the upper jaw of a fish. Yet, developmentally, these bones are the same thing. The stapes is a second-arch bone, as is the corresponding bone in a shark and a fish—the hyomandibula. But the hyomandibula is not an ear bone; recall that fish and sharks do not have ears. In our aquatic cousins, this bone is a large rod that connects the upper jaw to the braincase. Despite the apparent differences in the function and shape of these bones, the similarities between the hyomandibula and the stapes extend even to the nerves that supply them. The key nerve for the functioning of both bones is the second-arch nerve, the facial nerve. We thus have a situation where two very different bones have similar developmental origins and patterns of innervation. Is there an explanation for this?
Again, we look to the fossils. As we trace the hyomandibula from sharks to creatures like
Tiktaalik
to amphibians, we can see how it gets smaller and smaller, ultimately shifting position from the upper jaw to play a role in hearing. The name changes, too. When it is big and supporting the jaw, we call it a hyomandibula; when it is small and functions in hearing, it is known as a stapes. This shift happened when the descendants of fish began to walk on land. Hearing in water is different from hearing on land, and the small size and position of the stapes makes it ideal for picking up vibrations in air. The new ability came about by modifying the upper jawbone of a fish.
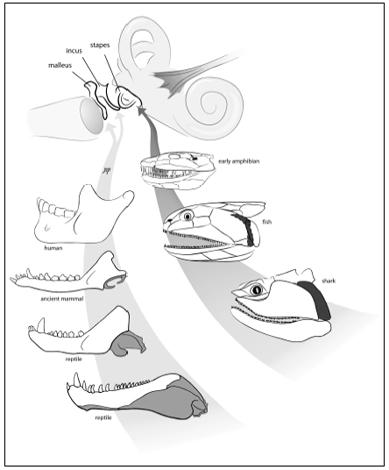
We can trace bones from gill arches to our ears, first during the transition from fish to amphibian (right), and later during the shift from reptile to mammal (left).
Our middle ear contains a record of two of the great transformations in the history of life. The origin of our stapes, and its transformation from a jaw support bone to an ear bone, began when fish started to walk on land. The other big event took place during the origin of mammals, when bones at the back of a reptile jaw became our malleus and incus.
Now let’s go further inside the ear—to the inner ear.
THE INNER EAR—GELS MOVING AND HAIRS BENDING

Move through the external ear, go deeper inside, past the eardrum and three middle ear bones, and you end up deep inside the skull. Here you will find the inner ear—tubes and some gel-filled sacs. In humans, as in other mammals, the bony tubes take the snail-shell shape that is so strikingly apparent in the anatomy lab.
The inner ear has different parts dedicated to different functions. One part is used in hearing, another in telling us which way our head is tilted, and still another in recording how fast our head is accelerating or stopping. In carrying out each of these functions, the inner ear works in roughly the same way.
The several parts of the inner ear are filled with a gel that can move. Specialized nerve cells send hairlike projections into this gel. When the gel moves, the hairs on the ends of the nerve cells bend. When these hairs bend, the nerve cells send an electrical impulse to the brain, where it is recorded as sound, position, or acceleration.
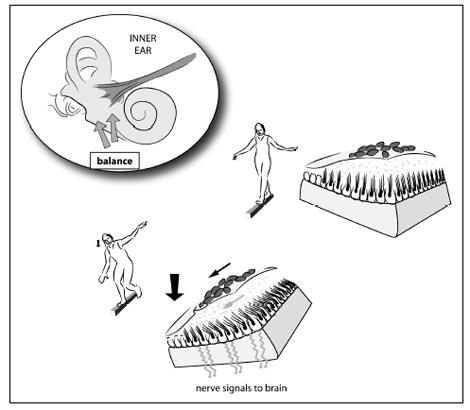
Each time you tilt your head, the tiny rocks on the fluid-filled sacs move. In doing so, they bend nerve endings inside the sacs and cause an impulse to be sent to your brain saying “Your head is tilted.”
To envision the structure that tells us where our head is in space, imagine a Statue of Liberty snow globe. The snow globe is made of plastic and filled with gel. When you shake it, the gel moves and the “snow” falls on Lady Liberty. Now imagine a snow globe made of a flexible membrane. Pick it up and tilt it, and the whole thing will flop about, causing the gel inside to swish around. This, on a much smaller scale, is what we have inside our ears. When we bend our heads, these contraptions flop around, causing the usual chain of events: the gel inside swishes, the hair projections on the nerves bend, and an impulse is sent back to our brains.
In us, this whole system is made even more sensitive by the presence of tiny rock-like structures on top of the membrane. As we bend our heads, the rocks accentuate the flopping of the membrane, causing the gel to move even more. This increases the sensitivity of the system, enabling us to perceive small differences in position. Tilt your head, and little rocks inside your skull move.
You can probably imagine how tough it would be to live in outer space. Our sensors are tuned to work in the earth’s gravity, not in a gravity-free space capsule. Floating around, our eyes recording one version of up and down, our inner ear sensors totally confused, it is all too easy to get sick. Space sickness has been a real problem for these very reasons.
The way we perceive acceleration is based on yet another part of our inner ear, connected to the previous two. There are three gel-filled tubes inside the ear; each time we accelerate or stop, the gel inside the tubes moves, causing the nerve cells to bend and stimulate a current.
The whole system we use to perceive position and acceleration is connected to our eye muscles. The motion of our eyes is controlled by eight small muscles attached to the side walls of the eyeball. The muscles contract to move the eye up, down, left, and right. We can move our eyes voluntarily by contracting these muscles each time we decide to look in a new direction; but some of the most fascinating properties of these muscles relate to their involuntary action. They move our eyes all the time, without our even thinking about it.
To appreciate the sensitivity of this eye-muscle link, move your head back and forth while looking at the page. Keep your eyes fixed in one place as you move your head.
What happened during this experiment? Your eyes stayed fixed on a single point while your head moved. This motion is so commonplace that we take it for granted, but it is incredibly complex. Each of the eight muscles in both eyes is responding to the movement of the head. Sensors in your head, which I’ll describe in the next section, record the direction and velocity of your head’s movement. These signals are carried to the brain, which then sends out signals telling your eye muscles to fire. Think about that the next time you fix your gaze as your head is moving. This system can misfire, and misfires have much to tell us about our general well-being.
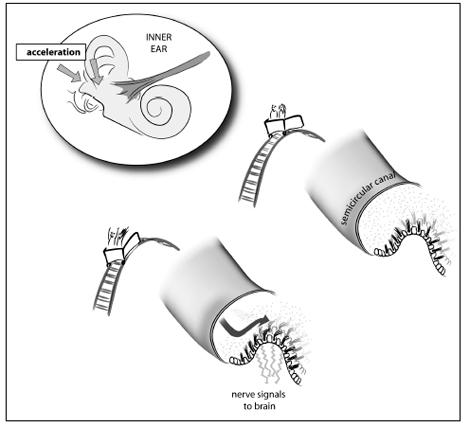
Every time we accelerate, fluid in the inner ear swishes. The swish is transformed into a nerve impulse that is sent to the brain.
An easy way to understand the inner ear–eye connection is to interfere with it. One way humans do this is to imbibe too much alcohol. Drinking too much ethanol leads us to do silly things because our inhibitions are lowered. Drinking
way
too much gives us the spins. And the spins often predict a lousy morning ahead, hungover, with more spins, nausea, and headache.
When we drink too much, we are putting lots of ethanol into our bloodstream, but the fluid inside our ear tubes initially contains very little. As time passes, however, the alcohol diffuses from our blood into the gel of the inner ear. Alcohol is lighter than the gel, so the result of the diffusion is like the result of pouring alcohol into a glass of olive oil. Just as the oil moves around in the glass as the alcohol enters, so the gel inside our ear swirls. The convection wreaks havoc on the intemperate among us. Our hair cells are stimulated and our brain thinks we are moving. But we are not moving; we are slumped in a corner or hunched on a barstool. Our brain has been tricked.
The problem extends to our eyes. Our brain thinks we are spinning, and it passes this information to our eye muscles. The eyes twitch in one direction (usually to the right) as we try to track an object moving from side to side. If you prop open the eyes of someone who is stone drunk, you might see this stereotypical twitch, called nystagmus. Police know this well, and often look for nystagmus in people whom they have stopped for driving erratically.
Massive hangovers involve a slightly different response. The day after the binge, your liver has done a remarkably efficient job of removing the alcohol from your bloodstream. Too efficient, for we still have alcohol in the tubes in our ears. That alcohol then diffuses from the gel back into the bloodstream, and in doing so it once more sets the gel in motion: the spins again. Take the same heavy drinker whose eyes you saw twitch to the right the night before and look at him during the hangover. His eyes might still twitch, but in the opposite direction.