Why Evolution Is True (8 page)
Read Why Evolution Is True Online
Authors: Jerry A. Coyne

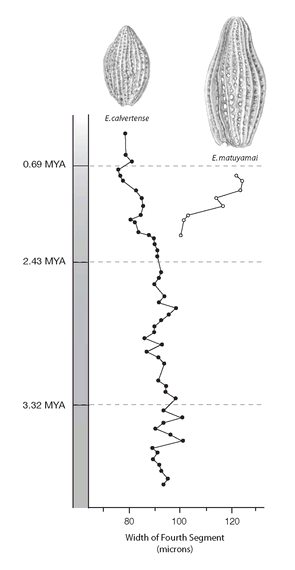
FIGURE 7
. Evolution and speciation in two species of the planktonic radiolarian
Eucyrtidium,
taken from a sediment core spanning more than 3.5 million years. The points represent the width of the fourth segment, shown as the average of each species at each section of the core. In areas to the north of where this core was taken, an ancestral population of E.
calvertense
became larger, gradually acquiring the name E.
matuyamai
as it became larger. E.
matuyamai
then reinvaded the range of its relative, as shown on the graph, and both species, now living in the same place, began to diverge in body size. This divergence may have been the result of natural selection acting to reduce competition for food between the two species.
. Evolution and speciation in two species of the planktonic radiolarian
Eucyrtidium,
taken from a sediment core spanning more than 3.5 million years. The points represent the width of the fourth segment, shown as the average of each species at each section of the core. In areas to the north of where this core was taken, an ancestral population of E.
calvertense
became larger, gradually acquiring the name E.
matuyamai
as it became larger. E.
matuyamai
then reinvaded the range of its relative, as shown on the graph, and both species, now living in the same place, began to diverge in body size. This divergence may have been the result of natural selection acting to reduce competition for food between the two species.
But what counts as fossil evidence for a major evolutionary transition? According to evolutionary theory, for every two species, however different, there was once a single species that was the ancestor of both. We could call this one species the “missing link.” As we’ve seen, the chance of finding that single ancestral species in the fossil record is almost zero. The fossil record is simply too spotty to expect that.
But we needn’t give up, for we can find some
other
species in the fossil record, close cousins to the actual “missing link,” that document common ancestry equally well. Let’s take one example. In Darwin’s day, biologists conjectured from anatomical evidence, such as similarities in the structure of hearts and skulls, that birds were closely related to reptiles. They speculated that there must have been a common ancestor that, through a speciation event, produced two lineages, one eventually yielding all modern birds and the other all modern reptiles.
other
species in the fossil record, close cousins to the actual “missing link,” that document common ancestry equally well. Let’s take one example. In Darwin’s day, biologists conjectured from anatomical evidence, such as similarities in the structure of hearts and skulls, that birds were closely related to reptiles. They speculated that there must have been a common ancestor that, through a speciation event, produced two lineages, one eventually yielding all modern birds and the other all modern reptiles.
What would this common ancestor have looked like? Our intuition is to say that it would have resembled something halfway between a modern reptile and a modern bird, showing a mixture of features from both types of animal. But this need not be the case, as Darwin clearly saw in
The Origin:
The Origin:
I have found it difficult, when looking at any two species, to avoid picturing to myself, forms directly intermediate between them. But this is a wholly false view; we should always look for forms intermediate between each species and a common but unknown progenitor; and the progenitor will generally have differed in some respects from all of its modified descendants.
Because reptiles appear in the fossil record before birds, we can guess that the common ancestor of birds and reptiles was an
ancient reptile,
and would have looked like one. We now know that this common ancestor was a dinosaur. Its overall appearance would give few clues that it was indeed a “missing link”—that one lineage of descendants would later give rise to all modern birds, and the other to more dinosaurs. Truly birdlike traits, such as wings and a large breastbone for anchoring the flight muscles, would have evolved only later on the branch leading to birds. And as that lineage itself progressed from reptiles to birds, it sprouted off many species having mixtures of reptilelike and birdlike traits. Some of those species went extinct, while others continued evolving into what are now modern birds. It is to these groups of ancient species, the relatives of species near the branch point, that we must look for evidence of common ancestry.
ancient reptile,
and would have looked like one. We now know that this common ancestor was a dinosaur. Its overall appearance would give few clues that it was indeed a “missing link”—that one lineage of descendants would later give rise to all modern birds, and the other to more dinosaurs. Truly birdlike traits, such as wings and a large breastbone for anchoring the flight muscles, would have evolved only later on the branch leading to birds. And as that lineage itself progressed from reptiles to birds, it sprouted off many species having mixtures of reptilelike and birdlike traits. Some of those species went extinct, while others continued evolving into what are now modern birds. It is to these groups of ancient species, the relatives of species near the branch point, that we must look for evidence of common ancestry.
Showing common ancestry of two groups, then, does not require that we produce fossils of the precise single species that was their common ancestor, or even species on the direct line of descent from an ancestor to descendant. Rather, we need only produce fossils having the types of traits that link two groups together, and, importantly, we must also have the dating evidence showing that those fossils occur at the right time in the geological record. A “transitional species” is not equivalent to “an ancestral species”; it is simply a species showing a mixture of traits from organisms that lived both before and after it. Given the patchiness of the fossil record, finding these forms at the proper times in the record is a sound and realistic goal. In the reptile-to-bird transition, for instance, the transitional forms should look like early reptiles, but with some birdlike traits. And we should find these transitional fossils after reptiles had already evolved, but before modern birds appeared. Further, transitional forms don’t have to be on the direct line of descent from an ancestor to a living descendant—they could be evolutionary cousins that went extinct. As we’ll see, the dinosaurs that gave rise to birds sported feathers, but some feathered dinosaurs continued to persist well after more birdlike creatures had evolved. Those later feathered dinosaurs still provide evidence for evolution, because they tell us something about where birds came from.
The dating and—to some extent—the physical appearance of transitional creatures, then, can be predicted from evolutionary theory. Some of the more recent and dramatic predictions that have been fulfilled involve our own group, the vertebrates.
Onto the Land: From Fish to AmphibiansOne of the greatest fulfilled predictions of evolutionary biology is the discovery, in 2004, of a transitional form between fish and amphibians. This is the fossil species
Tiktaalik
roseae, which tells us a lot about how vertebrates came to live on the land. Its discovery is a stunning vindication of the theory of evolution.
Tiktaalik
roseae, which tells us a lot about how vertebrates came to live on the land. Its discovery is a stunning vindication of the theory of evolution.
Until about 390 million years ago, the only vertebrates were fish. But, 30 million years later, we find creatures that are clearly
tetrapods:
four-footed vertebrates that walked on land. These early tetrapods were like modern amphibians in several ways: they had flat heads and bodies, a distinct neck, and well-developed legs and limb girdles. Yet they also show strong links with earlier fishes, particularly the group known as “lobe-finned fishes,” so called because of their large bony fins that enabled them to prop themselves up on the bottom of shallow lakes or streams. The fishlike structures of early tetrapods include scales, limb bones, and head bones (figure 8).
tetrapods:
four-footed vertebrates that walked on land. These early tetrapods were like modern amphibians in several ways: they had flat heads and bodies, a distinct neck, and well-developed legs and limb girdles. Yet they also show strong links with earlier fishes, particularly the group known as “lobe-finned fishes,” so called because of their large bony fins that enabled them to prop themselves up on the bottom of shallow lakes or streams. The fishlike structures of early tetrapods include scales, limb bones, and head bones (figure 8).
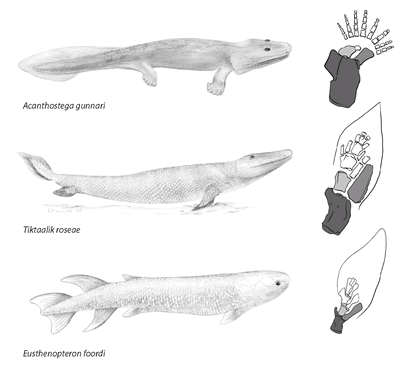
FIGURE 8
. Invasion of the land. An early lobe-finned fish (
Eusthenopteron foordi
) from about 385 million years ago; a land-dwelling tetrapod (
Acanthostega gunnari
) from Greenland, about 365 million years ago; and the transitional form,
Tiktaalik
roseae, from Ellesmere Island, about 375 million years ago. The intermediacy of
Tiktaalik’s
body form is mirrored by the intermediacy of its limbs, which have a bone structure in between that of the sturdy fins of the lobe-finned fish and the even sturdier walking limbs of the tetrapod. Shaded bones are those that evolved into the arm bones of modern mammals: the bone with darkest shading will become our humerus, and the medium- and light-shaded bones will become the radius and ulna, respectively.
. Invasion of the land. An early lobe-finned fish (
Eusthenopteron foordi
) from about 385 million years ago; a land-dwelling tetrapod (
Acanthostega gunnari
) from Greenland, about 365 million years ago; and the transitional form,
Tiktaalik
roseae, from Ellesmere Island, about 375 million years ago. The intermediacy of
Tiktaalik’s
body form is mirrored by the intermediacy of its limbs, which have a bone structure in between that of the sturdy fins of the lobe-finned fish and the even sturdier walking limbs of the tetrapod. Shaded bones are those that evolved into the arm bones of modern mammals: the bone with darkest shading will become our humerus, and the medium- and light-shaded bones will become the radius and ulna, respectively.
How did early fish evolve to survive on land? This was the question that interested—or rather obsessed—my University of Chicago colleague Neil Shubin. Neil had spent years studying the evolution of limbs from fins, and was driven to understand the earliest stages of that evolution.
This is where the prediction comes in. If there were lobe-finned fishes but no terrestrial vertebrates 390 million years ago, and clearly terrestrial vertebrates 360 million years ago, where would you expect to find the transitional forms? Somewhere in between. Following this logic, Shubin predicted that if transitional forms existed, their fossils would be found in strata around 375 million years old. Moreover, the rocks would have to be from freshwater rather than marine sediments, because late lobe-finned fish and early amphibians both lived in fresh water.
Searching his college geology textbook for a map of exposed freshwater sediments of the right age, Shubin and his colleagues zeroed in on a paleontologically unexplored region of the Canadian Arctic: Ellesmere Island, which sits in the Arctic Ocean north of Canada. And after five long years of fruitless and expensive searching, they finally hit pay dirt: a group of fossil skeletons stacked one atop another in sedimentary rock from an ancient stream. When Shubin first saw the fossil face poking out of the rock, he knew that he had at last found his transitional form. In honor of the local Inuit people and the donor who helped fund the expeditions, the fossil was named
Tiktaalik roseae
(“Tiktaalik” means “large freshwater fish” in Inuit, and “roseae” is a cryptic reference to the anonymous donor).
Tiktaalik roseae
(“Tiktaalik” means “large freshwater fish” in Inuit, and “roseae” is a cryptic reference to the anonymous donor).
Tiktaalik
has features that make it a direct link between the earlier lobe- finned fish and the later amphibians (figure 8). With gills, scales, and fins, it was clearly a fish that lived its life in water. But it also has amphibianlike features. For one thing, its head is flattened like that of a salamander, with the eyes and nostrils on top rather than on the sides of the skull. This suggests that it lived in shallow water and could peer, and probably breathe, above the surface. The fins had become more robust, allowing the animal to flex itself upward to help survey its surroundings. And, like the early amphibians,
Tiktaalik
has a neck. Fish don’t have necks—their skull joins directly to their shoulders.
has features that make it a direct link between the earlier lobe- finned fish and the later amphibians (figure 8). With gills, scales, and fins, it was clearly a fish that lived its life in water. But it also has amphibianlike features. For one thing, its head is flattened like that of a salamander, with the eyes and nostrils on top rather than on the sides of the skull. This suggests that it lived in shallow water and could peer, and probably breathe, above the surface. The fins had become more robust, allowing the animal to flex itself upward to help survey its surroundings. And, like the early amphibians,
Tiktaalik
has a neck. Fish don’t have necks—their skull joins directly to their shoulders.
Most important,
Tiktaalik
has two novel traits that were to prove useful in helping its descendants invade the land. The first is a set of sturdy ribs that helped the animal pump air into its lungs and move oxygen from its gills
(Tik
taalik could breathe both ways). And instead of the many tiny bones in the fins of lobe-finned fish,
Tiktaalik
had fewer and sturdier bones in the limbs—bones similar in number and position to those of every land creature that came later, including ourselves. In fact, its limbs are best described as part fin, part leg.
Tiktaalik
has two novel traits that were to prove useful in helping its descendants invade the land. The first is a set of sturdy ribs that helped the animal pump air into its lungs and move oxygen from its gills
(Tik
taalik could breathe both ways). And instead of the many tiny bones in the fins of lobe-finned fish,
Tiktaalik
had fewer and sturdier bones in the limbs—bones similar in number and position to those of every land creature that came later, including ourselves. In fact, its limbs are best described as part fin, part leg.
Clearly Tiktaalik was well adapted to live and crawl about in shallow waters, peek above the surface, and breathe air. Given its structure, we can envision the next, critical evolutionary step, which probably involved a novel behavior. A few of
Tiktaalik’s
descendants were bold enough to venture out of the water on their sturdy fin-limbs, perhaps to make their way to another stream (as the bizarre mudskipper fish of the tropics does today), to avoid predators, or perhaps to find food among the many giant insects that had already evolved. If there were advantages to venturing onto land, natural selection could mold those explorers from fish into amphibians. That first small step ashore proved a great leap for vertebrate-kind, ultimately leading to the evolution of every land-dwelling creature with a backbone.
Tiktaalik’s
descendants were bold enough to venture out of the water on their sturdy fin-limbs, perhaps to make their way to another stream (as the bizarre mudskipper fish of the tropics does today), to avoid predators, or perhaps to find food among the many giant insects that had already evolved. If there were advantages to venturing onto land, natural selection could mold those explorers from fish into amphibians. That first small step ashore proved a great leap for vertebrate-kind, ultimately leading to the evolution of every land-dwelling creature with a backbone.
Tiktaalik
itself was not ready for life ashore. For one thing, it had not yet evolved a limb that would allow it to walk. And it still had internal gills for breathing underwater. So we can make another prediction. Somewhere, in freshwater sediments about 380 million years old, we’ll find a very early land-dweller with reduced gills and limbs a bit sturdier than those of
Tiktaalik.
itself was not ready for life ashore. For one thing, it had not yet evolved a limb that would allow it to walk. And it still had internal gills for breathing underwater. So we can make another prediction. Somewhere, in freshwater sediments about 380 million years old, we’ll find a very early land-dweller with reduced gills and limbs a bit sturdier than those of
Tiktaalik.
Tiktaalik shows that our ancestors were flat-headed predatory fish who lurked in the shallow waters of streams. It is a fossil that marvelously connects fish with amphibians. And equally marvelous is that its discovery was not only anticipated, but predicted to occur in rocks of a certain age and in a certain place.
The best way to experience the drama of evolution is to see the fossils for yourself, or better yet, handle them. My students had this chance when Neil brought a cast of
Tiktaalik
to class, passed it around, and showed how it filled the bill of a true transitional form. This was, to them, the most tangible evidence that evolution was true. How often do you get to put your hands on a piece of evolutionary history, much less one that might have been your distant ancestor?
Into Thin Air: The Origin of BirdsTiktaalik
to class, passed it around, and showed how it filled the bill of a true transitional form. This was, to them, the most tangible evidence that evolution was true. How often do you get to put your hands on a piece of evolutionary history, much less one that might have been your distant ancestor?
Of what use is half a wing? Ever since Darwin, that question has been raised to cast doubt on evolution and natural selection. Biologists tell us that birds evolved from early reptiles, but how could a land-dwelling animal evolve the ability to fly? Natural selection, creationists argue, could not explain this transition, because it would require intermediate stages in which animals have just the rudiments of a wing. This would seem more likely to encumber a creature than to give it a selective advantage.
Other books
The Virgin Blue by Tracy Chevalier
Sweets for Sale: A Rough Stranger Sex Erotica Story by Sanders, Amy
Poison by Chris Wooding
Love on Lavender Island (A Lavender Island Novel Book 2) by Lauren Christopher
Hervey 11 - On His Majesty's Service by Allan Mallinson
High Wild Desert by Ralph Cotton
Legacy of Lies by Elizabeth Chandler
Island of the Forbidden by Hunter Shea
High and Dry by Sarah Skilton
The Nightmare by Lars Kepler