The Act of Creation (61 page)
Read The Act of Creation Online
Authors: Arthur Koestler

In an earlier paper (1941 a, p. 23) Weiss distinguished the following
levels of the hierarchy:
it one step further downward in the hierarchy. Even the lowest among
the six levels is a very complex affair. The individual muscle cell of a
striped muscle -- usually called a muscle fibre -- is a long, cylindrical
structure surrounded by a membrane. Its principal functions are: (a)
to serve as a receptor for nerve impulses which reach it at the synapse
through a chemical transmitter; (b) to re-code this message into an
electro-chemical excitation spreading along its surface; (c) to relay
it to the actual contractile structure, and to provide the energy for
the contraction in the cell's internal energy currency -- ATP; (d) to
contract.* This involves at least three distinct processes (acetylcholine
transmission; sodium-potassium action potentials; activation by ATP)
entering successively into action between the synaptic and the filament
levels, with mechanical contraction as the end product. Thus the ultimate
'motor unit' at the base of the hierarchy is not the cell itself, but
the apparatus within it which provides the contraction.
That apparatus is, broadly speaking, a kind of cylindrical cable,
the fibre, composed of a bundle of fibrils, each in turn composed of
bundles of filaments. The filaments are of two varieties, a thick and
a thin one, the former supposed to be consisting of molecular threads
of actin, the second of myosin. The combination of these two proteins --
acto-myosin -- is a substance which contracts when activated by the energy
carrier of the cell, ATP. The mechanism of the contraction is presumed
to be a telescoping into each other of the thick and fine bundles of
filaments. [7*]
It is assumed, then, that muscle movement is due to the chemical action
of ATP on contractile proteins; and moreover that essentially the same
process is responsible for the streaming motion of amoeba, the rowing
motion of cilia, and the tail-stroke motion of flagella. Amoeba,
as I have mentioned before (note to p. 423, previous chapter) are
capable of changing from amoeboid to flagellate form -- and back. Thus
at the very bottom of the hierarchy we find the same basic, universal
mechanism -- the archetype of organic motility as it were -- throughout
the whole animal kingdom from amoeba to man; and within man, we again
see it at work, serving such varied functions as the swimming of his
spermatozoa, the bristling of his hair, and the flexing of his muscles
in a tennis-stroke. It is a mechanism or apparatus with a high degree
of autonomy -- and it reminds one of the equally autonomous functioning
and universal occurrence of the power-plant-organelle, the mitochondria.
However close we seem to get to rock-bottom in the organic hierarchy,
we find complex, integrated sub-wholes leading a relatively autonomous
existence. Viruses have been compared to 'nomadic' genes (though
'freelancing' genes might be a more appropriate description).
Even a dead muscle cell (dissected and soaked in a cold glycerine solution
for months, which makes it very dead indeed) will contract when exposed to
the chemical trigger-action of ATP. The glycerine destroys cell-components
serving its higher functions, but the essential structure of the fibrils
-- the structural matrix of the basic motor unit -- remains intact; and
it is still capable of functioning according to its built-in code. This
function is of course a fixed reaction, as one would expect on the lowest
level of the hierarchy; it is activated by the trigger-action of ATP on
the acto-myosin. The environment of this matrix is represented by the
temperature, oxygen supply, degree of fatigue in the cell, but the degrees
of freedom of the fibre to adjust to these conditions boil down to an
'all-or-nothing' strategy: the alternative is to twitch or not to twitch.
On the higher levels of the hierarchy, the autonomous function-patterns of
muscles and muscle-complexes are even more in evidence; at the same time
the degrees of freedom in the matrix allowing for adaptable performance
increase with each level. Muscles dissected from the body and put into
Ringer solution will contract normally for hours. Practically any part
of an animal's heart, a muscular strip, and even a single muscle cell
grown on a blood dot, will continue to go on beating in its own intrinsic
rhythm. The heart of the chick embryo starts beating before any nerve
cells have grown into it, and the hearts of frogs and tortoises will
go on beating normally if the nerve supply is cut. Some smooth muscles
equally show a rhythm of their own.
But complementary to this
Eigenfunktion
, or functional autonomy,
is control by a centre or system on the next higher level. The heart has
its own pacemaker-system which is in itself a threefold hierarchy; under
normal conditions, the sino-auricular node, the fastest part, acts as a
pacemaker; but if it is prevented from doing so, the auro-ventricular
node will take over, and as a last resort (in experiments carried out
on frog and tortoise) yet a third centre may enter into action. The
pacemaker-system is, in its turn, subject to regulatory control by
sympathetic and parasympathetic nerves and by hormones, which will
speed up or slow down the rate of beat by order of centres in the
hypothalamus. Other organs -- kidney, intestine, stomach -- also have
their self-regulating, intrinsic codes which assure their status as
autonomous subwholes, while at the same time they function as parts
in a multi-levelled hierarchy. Even the mid-brain centres which control
temperature, metabolism, food and liquid intake, respiration, etc. -- even
these homeostatic controls responsible for maintaining the equilibrium
of the
milieu intérieur
, turn out to be subject to the
control of still higher levels. They are 'biassed homeostats' which can be
'set', as a thermostat is 'set' by the tenant to keep a higher or lower
room temperature. [8] And so the top of the hierarchy which controls
the controls recedes into a cloud -- just as its base is embedded in
the murky problem of what constitutes living matter -- and ultimately
dissolves into genetic mutations with thresholds on the quantum level.
The Goldfish and the Crab
Getting back to earth, that is, to the medium levels of the hierarchy
-- the levels 3, 4, and 5 in Weiss's schema -- we find, fortunately,
more precise indications about its manner of working.
Von Holst's study of the swimming motions of fish revealed a distinct
three-step hierarchy: (a) the motions of the rays within a single fin,
due to the alternative contractions of two antagonistic pairs of muscles;
(b) the motions of the fin as a whole; and (c) the co-ordination
of the motions of all the fins. In the anaesthetized goldfish, the
swinging motions of each individual ray remain perfectly regular,
but their co-ordination within the fin is disturbed: they flutter in
disorder. The anaesthesia evidently does not affect the integrative
centres on the lowest (a) level, but puts the higher nervous centres
on the (b) level out of action. On the next higher, (c) level, the
pectoral fin acts as a kind of pacemaker by imposing, or superimposing,
its own rhythm on the caudal fins -- the so-called 'magnet effect'. This
whole locomotor hierarchy is relatively independent of sensory stimuli,
for fishes and tadpoles go on swimming in perfect co-ordination if they
have been disafferentated, i.e. if all the main sensory connections have
been severed. Von Hoist concludes that the stimulus-response schema does
not apply to the autonomous locomotive hierarchy, and that 'the reflex
is not the primary element of behaviour but a device for adapting the
primary automatism to changing peripheral conditions'. [9]
Higher up on the evolutionary ladder we find increasing flexibility of
motor skills. In a series of famous experiments, von Buddenbrook and
Bethe have shown that the removal of one or several legs from centipedes,
spiders, and other insects does not lead to disorganization, but to a
spontaneous rearrangement of the whole pattern of locomotion which is
instantaneous and not preceded by trial-and-error learning. The normal
progress of an insect or crab is the so-called 'cross-amble'. If 'L' and
'R' stand for left and right, and the index numbers stand for the order
of legs from front to rear, the crab's locomotive code is as follows: R1,
L2, R3, L4, R5, etc., are stepping simultaneously; then L1, R2, L3, R4,
L5 -- are stepping simultaneously; and so on. If, now, the left front
leg is removed, the pattern changes instantaneously to: R1, L3, R3, L5,
etc.; followed by L2, R2, L4, R4, L6, etc. The crab's progress before
and after loss of the left front leg:
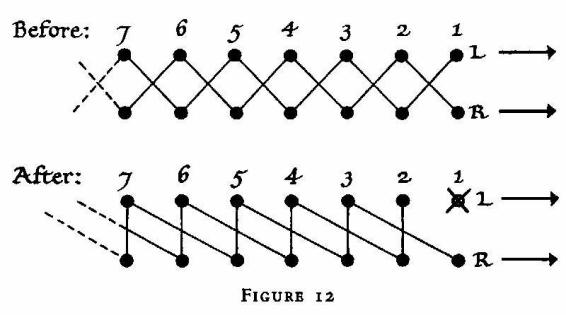 This transformation indicates that the front legs act as 'pacemakers';
This transformation indicates that the front legs act as 'pacemakers';
this makes it impossible for the animal to adopt the simpler solution
of preserving the original pattern minus L1: for in this case R2 would
become the second pacemaker and both pacemakers would be on the right side
of the animal. The crab's behaviour provides us with a rather elegant
example how a motor skill can be adapted to changed conditions while
preserving the basic pattern laid down in its code. I have mentioned
other examples earlier on -- from the spider's net to the pianist who
transposes a tune from one key to another. The experiments to be described
presently illustrate the challenging nature of the problem.
Shuffling the Salamander's Limbs
Weiss's transplantation experiments date back to the 1920s and proved to
be, as one author said, 'of immeasurable positive significance for the
appraisal of centro-peripheral co-ordination in nervous function'. [10]
Weiss grafted fully developed limbs of salamander as super-numeraries
on to normal animals which thus had five limbs instead of four. The
additional limb was always grafted next to a normal one, and in
the process some of the nerve-fibres supplying the normal limb were
severed. At first the transplant limb hung inertly from the body as
a mere appendage -- the fifth wheel of the cart. However, after a few
weeks, it began to give signs of movement, and within a short time it
functioned in complete synchronicity with the adjacent normal limb,
as its equal in vigour and co-ordination.
The implications of this phenomenon were described by Weiss as follows
(italics in the original):
 Assuming D0 and E0 to be a pair of antagonistic muscles -- how can they
Assuming D0 and E0 to be a pair of antagonistic muscles -- how can they
properly function if both now depend on the same nerve supply? And what
about At, Bt, Ct? Yet this 'boundless confusion of conduction paths'
nevertheless produces perfectly coordinated movement. Weiss concludes
that it is not the topographical layout of the pathways which matters,
but the specific properties of the excitation transmitted by them; in
other words, that although each muscle of the added limb will receive
a chaotic medley of excitations, it will respond selectively to such
excitations only which are appropriate to it:
analogy of selective resonance in a broad sense. The acoustic analysers
of the ear each respond to one particular pitch and to one only, thus
analysing a complex clang into in harmonic elements. Mutatis mutandis,
Weiss assumes that:
is not contradicted by the indiscriminate responses of muscles to
electro-galvanic stimulation. The latter is an artificial, gross
stimulation which compares to natural stimulation like a violent
a-periodic blast to a specific clang. 'Just as, both with the clang
and with the blast, the substratum carrying the movement is always the
same, i.e. the air, so obviously the medium in which both the organized
and the unorganized nerve impulse run their course is always the same,
i.e. the conductive substance of the nerve fibre. But just as the clang
sets a definite selection of resonators vibrating, whereas a noise or
blast causes them to resound all at once and without an exception; so
also only the organized impulse, built up of specific impulse-tones, is
capable of bringing the coordinated selection of muscles into activity,
while 'the artificially induced, unorganized impulse, by contrast,
forces every muscle whatsoever which it reaches into function.' [14]
Let me translate the picture that emerges from the experimental evidence
into the terms of the present theory. The locomotor
matrix
on
level 4 of the hierarchy (
levels of the hierarchy:
1. The level of the individual motor unit.This is as far as the schema proposed by Weiss goes. Now let me extend
2. All the motor units belonging to one muscle.
3. Co-ordinated functions of muscular complexes relating to a
single joint.
4. Coordinated movements of a limb as a whole.
5. Coordinated movements of a number of locomotor organs resulting
in locomotion.
6. "The highest level common to all animals", the movements of
"the animal as a whole".
it one step further downward in the hierarchy. Even the lowest among
the six levels is a very complex affair. The individual muscle cell of a
striped muscle -- usually called a muscle fibre -- is a long, cylindrical
structure surrounded by a membrane. Its principal functions are: (a)
to serve as a receptor for nerve impulses which reach it at the synapse
through a chemical transmitter; (b) to re-code this message into an
electro-chemical excitation spreading along its surface; (c) to relay
it to the actual contractile structure, and to provide the energy for
the contraction in the cell's internal energy currency -- ATP; (d) to
contract.* This involves at least three distinct processes (acetylcholine
transmission; sodium-potassium action potentials; activation by ATP)
entering successively into action between the synaptic and the filament
levels, with mechanical contraction as the end product. Thus the ultimate
'motor unit' at the base of the hierarchy is not the cell itself, but
the apparatus within it which provides the contraction.
That apparatus is, broadly speaking, a kind of cylindrical cable,
the fibre, composed of a bundle of fibrils, each in turn composed of
bundles of filaments. The filaments are of two varieties, a thick and
a thin one, the former supposed to be consisting of molecular threads
of actin, the second of myosin. The combination of these two proteins --
acto-myosin -- is a substance which contracts when activated by the energy
carrier of the cell, ATP. The mechanism of the contraction is presumed
to be a telescoping into each other of the thick and fine bundles of
filaments. [7*]
It is assumed, then, that muscle movement is due to the chemical action
of ATP on contractile proteins; and moreover that essentially the same
process is responsible for the streaming motion of amoeba, the rowing
motion of cilia, and the tail-stroke motion of flagella. Amoeba,
as I have mentioned before (note to p. 423, previous chapter) are
capable of changing from amoeboid to flagellate form -- and back. Thus
at the very bottom of the hierarchy we find the same basic, universal
mechanism -- the archetype of organic motility as it were -- throughout
the whole animal kingdom from amoeba to man; and within man, we again
see it at work, serving such varied functions as the swimming of his
spermatozoa, the bristling of his hair, and the flexing of his muscles
in a tennis-stroke. It is a mechanism or apparatus with a high degree
of autonomy -- and it reminds one of the equally autonomous functioning
and universal occurrence of the power-plant-organelle, the mitochondria.
However close we seem to get to rock-bottom in the organic hierarchy,
we find complex, integrated sub-wholes leading a relatively autonomous
existence. Viruses have been compared to 'nomadic' genes (though
'freelancing' genes might be a more appropriate description).
Even a dead muscle cell (dissected and soaked in a cold glycerine solution
for months, which makes it very dead indeed) will contract when exposed to
the chemical trigger-action of ATP. The glycerine destroys cell-components
serving its higher functions, but the essential structure of the fibrils
-- the structural matrix of the basic motor unit -- remains intact; and
it is still capable of functioning according to its built-in code. This
function is of course a fixed reaction, as one would expect on the lowest
level of the hierarchy; it is activated by the trigger-action of ATP on
the acto-myosin. The environment of this matrix is represented by the
temperature, oxygen supply, degree of fatigue in the cell, but the degrees
of freedom of the fibre to adjust to these conditions boil down to an
'all-or-nothing' strategy: the alternative is to twitch or not to twitch.
On the higher levels of the hierarchy, the autonomous function-patterns of
muscles and muscle-complexes are even more in evidence; at the same time
the degrees of freedom in the matrix allowing for adaptable performance
increase with each level. Muscles dissected from the body and put into
Ringer solution will contract normally for hours. Practically any part
of an animal's heart, a muscular strip, and even a single muscle cell
grown on a blood dot, will continue to go on beating in its own intrinsic
rhythm. The heart of the chick embryo starts beating before any nerve
cells have grown into it, and the hearts of frogs and tortoises will
go on beating normally if the nerve supply is cut. Some smooth muscles
equally show a rhythm of their own.
But complementary to this
Eigenfunktion
, or functional autonomy,
is control by a centre or system on the next higher level. The heart has
its own pacemaker-system which is in itself a threefold hierarchy; under
normal conditions, the sino-auricular node, the fastest part, acts as a
pacemaker; but if it is prevented from doing so, the auro-ventricular
node will take over, and as a last resort (in experiments carried out
on frog and tortoise) yet a third centre may enter into action. The
pacemaker-system is, in its turn, subject to regulatory control by
sympathetic and parasympathetic nerves and by hormones, which will
speed up or slow down the rate of beat by order of centres in the
hypothalamus. Other organs -- kidney, intestine, stomach -- also have
their self-regulating, intrinsic codes which assure their status as
autonomous subwholes, while at the same time they function as parts
in a multi-levelled hierarchy. Even the mid-brain centres which control
temperature, metabolism, food and liquid intake, respiration, etc. -- even
these homeostatic controls responsible for maintaining the equilibrium
of the
milieu intérieur
, turn out to be subject to the
control of still higher levels. They are 'biassed homeostats' which can be
'set', as a thermostat is 'set' by the tenant to keep a higher or lower
room temperature. [8] And so the top of the hierarchy which controls
the controls recedes into a cloud -- just as its base is embedded in
the murky problem of what constitutes living matter -- and ultimately
dissolves into genetic mutations with thresholds on the quantum level.
The Goldfish and the Crab
Getting back to earth, that is, to the medium levels of the hierarchy
-- the levels 3, 4, and 5 in Weiss's schema -- we find, fortunately,
more precise indications about its manner of working.
Von Holst's study of the swimming motions of fish revealed a distinct
three-step hierarchy: (a) the motions of the rays within a single fin,
due to the alternative contractions of two antagonistic pairs of muscles;
(b) the motions of the fin as a whole; and (c) the co-ordination
of the motions of all the fins. In the anaesthetized goldfish, the
swinging motions of each individual ray remain perfectly regular,
but their co-ordination within the fin is disturbed: they flutter in
disorder. The anaesthesia evidently does not affect the integrative
centres on the lowest (a) level, but puts the higher nervous centres
on the (b) level out of action. On the next higher, (c) level, the
pectoral fin acts as a kind of pacemaker by imposing, or superimposing,
its own rhythm on the caudal fins -- the so-called 'magnet effect'. This
whole locomotor hierarchy is relatively independent of sensory stimuli,
for fishes and tadpoles go on swimming in perfect co-ordination if they
have been disafferentated, i.e. if all the main sensory connections have
been severed. Von Hoist concludes that the stimulus-response schema does
not apply to the autonomous locomotive hierarchy, and that 'the reflex
is not the primary element of behaviour but a device for adapting the
primary automatism to changing peripheral conditions'. [9]
Higher up on the evolutionary ladder we find increasing flexibility of
motor skills. In a series of famous experiments, von Buddenbrook and
Bethe have shown that the removal of one or several legs from centipedes,
spiders, and other insects does not lead to disorganization, but to a
spontaneous rearrangement of the whole pattern of locomotion which is
instantaneous and not preceded by trial-and-error learning. The normal
progress of an insect or crab is the so-called 'cross-amble'. If 'L' and
'R' stand for left and right, and the index numbers stand for the order
of legs from front to rear, the crab's locomotive code is as follows: R1,
L2, R3, L4, R5, etc., are stepping simultaneously; then L1, R2, L3, R4,
L5 -- are stepping simultaneously; and so on. If, now, the left front
leg is removed, the pattern changes instantaneously to: R1, L3, R3, L5,
etc.; followed by L2, R2, L4, R4, L6, etc. The crab's progress before
and after loss of the left front leg:
this makes it impossible for the animal to adopt the simpler solution
of preserving the original pattern minus L1: for in this case R2 would
become the second pacemaker and both pacemakers would be on the right side
of the animal. The crab's behaviour provides us with a rather elegant
example how a motor skill can be adapted to changed conditions while
preserving the basic pattern laid down in its code. I have mentioned
other examples earlier on -- from the spider's net to the pianist who
transposes a tune from one key to another. The experiments to be described
presently illustrate the challenging nature of the problem.
Shuffling the Salamander's Limbs
Weiss's transplantation experiments date back to the 1920s and proved to
be, as one author said, 'of immeasurable positive significance for the
appraisal of centro-peripheral co-ordination in nervous function'. [10]
Weiss grafted fully developed limbs of salamander as super-numeraries
on to normal animals which thus had five limbs instead of four. The
additional limb was always grafted next to a normal one, and in
the process some of the nerve-fibres supplying the normal limb were
severed. At first the transplant limb hung inertly from the body as
a mere appendage -- the fifth wheel of the cart. However, after a few
weeks, it began to give signs of movement, and within a short time it
functioned in complete synchronicity with the adjacent normal limb,
as its equal in vigour and co-ordination.
The implications of this phenomenon were described by Weiss as follows
(italics in the original):
As could be incontrovertibly gathered from the microscopical
(post-mortem) investigation and reconstruction of the course of the
nerves in the original limb and in the transplanted limb, this is
what took place. The severed nerve fibres had vigorously split
up in the scar at the place of grafting. The branches had pressed
forward, and some of them had eventually met the degenerated nerve
paths of the transplanted limb. As fortuitously as they were
located and distributed, they had penetrated into these and
so had reached the muscles . . . in the most extraordinary and
indiscriminate tangle. . . . Moreover, those few paths belonging to
the normal extremity which had also been previously cut (in order to
obtain severed nerve stumps capable of regeneration for the supply
of the grafted limb) these too were filled with fresh nerves. In the
end, therefore, the relatively small number of ganglion cells, which
originally led to a small, limited section of the musculature of the
normal extremity were now not only connected with this very section
of muscle again, but in addition with the entire musculature of the
grafted limb. . . . Thus not only have the ganglion cells involved to
serve a terminal area several times as large as before; and not only
have they to serve muscles altogether different from. the previous
ones . . . but above all the previous rule, that one ganglion cell had
connections with only one muscle, now becomes the exception. Instead
the rule is now a boundless confusion of conduction paths. [11]
properly function if both now depend on the same nerve supply? And what
about At, Bt, Ct? Yet this 'boundless confusion of conduction paths'
nevertheless produces perfectly coordinated movement. Weiss concludes
that it is not the topographical layout of the pathways which matters,
but the specific properties of the excitation transmitted by them; in
other words, that although each muscle of the added limb will receive
a chaotic medley of excitations, it will respond selectively to such
excitations only which are appropriate to it:
The means by which the central nervous system maintains concord withTo account for the specific selectivity of muscle response, Weiss uses the
each muscle individually, does not consist in separate conduction
paths. . . . If one and the same nerve cell has to supply excitation
to several organs simultaneously, but if under these circumstances
only one single route common to all these end-organs is at its
disposal . . . then it is logical to assume that the periphery is
so constituted that a control of its functioning in a coordinated
manner inheres in itself. . . . We require . . . a mechanism of
positive selectivity in the end organ, which must explain us why,
when two muscles in the same state are given, one of them enters into
function and the other does not, although both, being connected with
the same nerve cell, receive excitation equally. . . . The nature of
every muscle is such that it does not react to every excitation from
the centre, but only to excitation of a quite definite form
which is characteristic for it. [12]
analogy of selective resonance in a broad sense. The acoustic analysers
of the ear each respond to one particular pitch and to one only, thus
analysing a complex clang into in harmonic elements. Mutatis mutandis,
Weiss assumes that:
. . . the total impulse flowing towards a particular peripheralHe then proceeds to show that the theory of selective response
region from the central nervous system can, metaphorically speaking,
forthwith be designated as an "excitation clang". The "excitation clang"
is composed of "excitation tones" for the varying muscles which are to
be activated at a given moment, and hence is constantly fluctuating in
its composition. . . . The process now is as follows: at the very same
time, the same "excitation clang" flows through all the motor
root fibres (at least all those supplying a given functional area of
considerable extent) towards the periphery. It flows equally
through all the fibres as if it had been indiscriminately poured
into a canal system and were flooding all the channels. Thus it arrives
at all the muscles which are in any way whatever connected with the
centre. But when it gets to this point it is analysed. Every
muscle, in accordance with its constitution, selects the components
appropriate to it from those eventually arriving, and acts as if these
components alone had arrived. And thus, although the very same impulse
streams to all the muscles and across every available route, only that
combination of muscles comes into action -- as is now intelligible --
which the central nervous system has provided for. [13]
is not contradicted by the indiscriminate responses of muscles to
electro-galvanic stimulation. The latter is an artificial, gross
stimulation which compares to natural stimulation like a violent
a-periodic blast to a specific clang. 'Just as, both with the clang
and with the blast, the substratum carrying the movement is always the
same, i.e. the air, so obviously the medium in which both the organized
and the unorganized nerve impulse run their course is always the same,
i.e. the conductive substance of the nerve fibre. But just as the clang
sets a definite selection of resonators vibrating, whereas a noise or
blast causes them to resound all at once and without an exception; so
also only the organized impulse, built up of specific impulse-tones, is
capable of bringing the coordinated selection of muscles into activity,
while 'the artificially induced, unorganized impulse, by contrast,
forces every muscle whatsoever which it reaches into function.' [14]
Let me translate the picture that emerges from the experimental evidence
into the terms of the present theory. The locomotor
matrix
on
level 4 of the hierarchy (
Other books
Snare: Road Kill MC (A Novel) by Marata Eros
Don't Call Me Ishmael by Michael Gerard Bauer
Wagers of Sin: Time Scout II by Robert Asprin, Linda Evans
RUNAWAY by Christie Ridgway
A Countess of Convenience by Sarah Winn
The Solitary Man by Stephen Leather
Born to Darkness by Suzanne Brockmann
On the Prowl by Christine Warren
Hard to Hold On by Shanora Williams
Blood Redemption (Angel's Edge #3) by Keire, Vicki