The Act of Creation (60 page)
Read The Act of Creation Online
Authors: Arthur Koestler

On the level of the zygote, the cell-matrix consists of biochemical
gradients and organelles; it is a structural mosaic equipped with axial
polarity which under normal conditions predetermines the head and tail
region, the blastopores, etc., of the embryo; but it also has striking
regulative properties revealed by experimental manipulation. With
progressive differentiation the regulative properties of the cell
diminish, and its degrees of 'genetic freedom' freeze up. On the level
of the morphogenetic field we again find self-regulating properties --
half the field will still form a complete limb -- and a mosaic-matrix
of cell-populations. The autonomous, self-assertive character of
morphogenetic fields is manifested in grafting and centrifuging
experiments; their dependent, or part character by the trivial fact that
they are kept in their proper size and place in the normally growing
organism. This will sound less of a truism when we turn to the phenomena
of regeneration (Chapter III). We shall find regulative character and
mosaic character, autonomy and subservience, the self-assertion of
the part and its dependence on the whole to be complementary aspects
on every level of the hierarchy in normal development and behaviour;
but also, that under abnormal conditions this ceases to be the case,
and that the part may then assert itself at the expense of the whole,
with sometimes beneficial, mostly destructive, effects.*
NOTES
To
p. 418
. This is, of courre, not meant to
belittle the enormous advantages of sexual over asexual reproduction.
p. 419
. The three-letter 'dictionary', for
instance, is partly a dictionary of synonyms: there are 4^3 = 64 triplets,
but only 20 arnino acids, and many of the latter are represented by more
than one code syllable.
p. 420
. 'Respiration' is an approximate
term. The process is in fact oxidative phosphorylation.
p. 422
. In some, probably extreme cases, the
nuclear changes are even more drastic. The nucleus of the fertilized egg
of the gall-midge contains forty chromosomes, and in the course of the
first few divisions these are faithfully duplicated. But in the fifth
division, only eight sets of chromosomes in the soma cells duplicate in
the orthodox manner; the other thirty-two fail to do so and gradually
dissolve in the cytoplasm. The future germ calls, however, which have
previously been segregated from the rest of the eggs, do not participate
in the fateful fifth division and preserve their chromosome complement
intact. Thus the nuclei of all specialized body cells have only eight
chromosomes, whereas the germ cells have forty. Cf. Fischberg, M. and
Blackler, A. W. (1961).
p. 423
. In a paper read at the British
Association Meeting in August 1962, L. Wolpert suggested that
differentiation resulted from the single cell's tendency to stick on to
that part of the gastrula wall best suited for it (the idea seems to have
been originated by T. Gusthafson). At the same meeting E. N. Willmer showed
that changes in the salt balance of the surrounding medium made amoebae
change from amoeboid to flagellate form, reversibly -- the implication
being that chemical gradients played an important part in the early
stages of differentiation (
New Scientist
, No 303, 6.9.1962, p. 492).
p. 425
. During maturation in the higher species,
the two types of control overlap; and pre-set biological time-clocks
seem to exercise some influence throughout adult life.
p. 428
. In this necessarily simplified
discussion of morphogenetic processes I have made no mention of
cytoplasmic inheritance and other complicating factors, which do not
affect the basic argument of this book.
embryos is G. E. Coghill's monumental study of
ambystoma
, a larval
form of salamander). [1] Coghill published his results in a series
of papers spread over a period of twenty-five years, 1914-39. Since
they are surprisingly seldom quoted outside the technical literature --
presumably because they ran against the behaviourist Zeitgeist -- I must
briefly summarize his conclusions.
was that elementary, local reflexes arise first, and are chained together
at a later stage. Thus the segmental reflex arcs of the earthworm would
develop first, as independent units aligned in a series perpendicular
to its axis, and only later on would they become connected, like rings
hanging from a festoon string, by the spinal cord. Coghill's work showed
that the opposite is true. In the salamander, development starts with
the growth of the motor-tracts of the cord axially from head to tail;
then this central bundle sends out collateral branches into the segmental
muscles, coordinating their actions in primitive, unitary patterns; the
sensory neurons become functional only at a later stage, and the local
reflex-arcs come last, as segregations of 'partial patterns' out of the
'total pattern' which preceded them. The whole development is centrifugal:
the stem precedes the branches, spontaneous undifferentiated movements
involving the whole neuro-muscular apparatus precede differentiated
movement, total responses precede specialized local responses. To give
an example: when the limbs develop, their first movements are entirely
dependent on and synchronized with the movements of the trunk. Only
later on do the limbs begin to move independently; the same applies to
the motions of head, mouth, etc. The growth of the nervous system from
beginning to end is dominated by 'a totally integrated matrix, and not
a progressive integration of primarily individuated units'. The organism
is not a sum of its reflexes, but on the contrary 'the mechanism of the
total pattern is an essential component of the performance of the part,
i.e. the reflex'. The stimulus-response scheme cannot explain even
embryonic behaviour, because movements appear long before the motor
neurons of the reflex arc are connected with the sensory neurons. This
centrifugal mode of development means that the individual acts on its
environment before it reacts to its environment.
development of the individual has been carried, structural provision
has been found for the perpetuation of spontaneity, autonomy, or
initiative as a factor in its behaviour. Any theory of motivation,
therefore, that attributes this function wholly to the environment, is
grossly inadequate.' The idea that instincts are chained reflexes must
be abandoned; instincts represent 'total action patterns in response to
relatively general situations'. Comparing the embryonic development of
ambystoma with that of the human foetus, Coghill sums up:
principles at work which we have discussed before. The neural plate
starts as a primordium with multipotential cell-populations which
differentiate in a series of steps into the brain, the spinal cord,
and its substructures. The 'wiring diagram' of the organism has a
standardized pattern -- an invariant code; but transplant experiments
again show the great flexibility of the 'neurogenetic skill' which
realizes that pattern. If a limb-bud from a salamander embryo is
transplanted to another embryo's flank, outgrowing nerve-fibres locate
the bud and establish a normal nerve pattern. The bulb-shaped tips of the
outgrowing nerves are apparently guided by submicroscopic structures in
the cell-matrix of the growing bud -- at least according to the current
'contact guidance' theory.
I have called differentiation of structure and integration of function
complementary aspects of a unitary process. But the 'functions' of the
growing embryo are different from the 'functions' of the adult. It has
been shown that the limb-buds and wing buds of chick embryos develop
into almost normal legs and wings if nerves are prevented from entering
them. This does not mean, of course, that differentiation of structure
comes first, and integration of function later on, as a separate act. For
the function of the leg-bud is to grow -- not to walk. Growth is a
function controlled by the genetic code; when growth is completed and
the time has come to walk, the nervous system takes over control; and
if it fails to do so, the muscle tissues will degenerate, as denervated
adult muscles do.*
Locomotor Hierarchies
'Whatever the nature of organizing relations may be,' J. Needham wrote
in 1932, 'they form the central problem of biology, and biology will
be fruitful in the future only if this is recognized. The hierarchy
of relations, from the molecular structure of carbon compounds to the
equilibrium of species and ecological wholes, will perhaps be the leading
idea of the future.' [2]
This prophecy has not come true. The Gestalt school's over-emphasis on
'wholeness', and the behaviourists' over-emphasis on 'simple elementary
processes' -- the so-called S.-R. (stimulus-response) scheme -- created
a controversy based on a fallacious alternative, and prevented a true
appreciation of the multi-layered hierarchic order to be found in all
manifestations of life.
Yet the idea is of course by no means new; hierarchies in nervous function
were proposed by Herbert Spencer in the 1870s, and elaborated by Hughlings
Jackson, Sherrington, and others. [3] The hierarchical character of
skills was demonstrated in great detail by Bryan and Harter in their
study of telegraphy and in Book's study of touch-typing (see below,
pp. 544
ff.) at the turn of the century,
but neither S-R psychologists nor Gestaltists paid attention to
them. Woodger (1929) attempted a formalization, by means of symbolic
logic, of certain types of hierarchies ('divisional hierarchies', 'spatial
hierarchies', 'genetic hierarchies', etc.) which are of somewhat abstract
interest. Heidenhain (1923) [4] proposed a hierarchy of 'histo-systems'
which are 'encapsulated' into one another (e.g. neuro-fibriles, neurons,
nerve fibres). Bertalanffy (1952) tried to make a distinction between
'hierarchies of parts', 'hierarchies of processes', 'hierarchies of
centralization', etc. Tinbergen defined instinct as a hierarchically
organized nervous mechanism -- but his mechanism is fixed and rigid
(see below p. 478). A stimulating discussion of the subject can be found
in Miller, Galanter, and Pribram's remarkable essay on 'Plans and the
Structure of Behaviour' (1960).
'habit family hierarchy', for instance, means just that (the ordering
of a group of interchangeable responses according to their strength),
and is not a hierarchy at all in the sense in which the term is used in
this book. I have summarized what I meant by it in the chapter 'Partness
and Wholeness' (Book One, Chapter XIII). A hierarchy, in this sense, is
not like a row of organ pipes; it is like a tree, arborizing downward. The
structural or functional entities on each level are autonomous sub-wholes
of complex pattern, but are represented on the next higher level as
units. In every organic hierarchy, to paraphrase Gertrude Stein's
statement about the rose, 'a part is a whole is a part is a whole'.
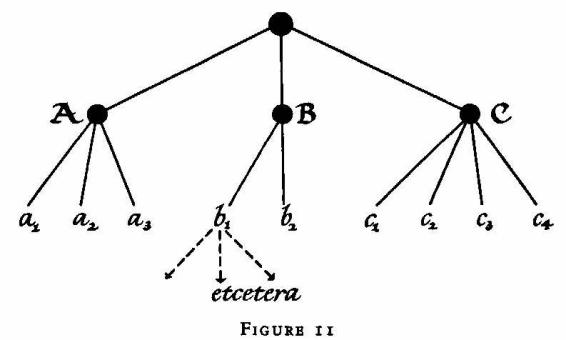 Perhaps the most satisfactory theoretical treatment of the concept of
Perhaps the most satisfactory theoretical treatment of the concept of
hierarchic order was given by Paul Weiss -- whose experimental work
was a major contribution towards providing the concept with a firm
empirical basis. The quotation which follows is from the celebrated Hixon
Symposium; its vivickss is enhanced by the fact that it is taken from an
ex tempore contribution by Weiss to the discussion of Lashley's paper on
'The Problem of Serial Order in Behaviour'* (my italics):
emerged since.
In the first place it has been found that intrinsic, rhythmic activity
of an autonomous character is not confined to motor nerves, but
that 'receptors also are spontaneously active even in the absence of
stimulation from environment.' [6] This spontaneous receptor activity,
while modified by environmental events, is under efferent control from
the central nervous system. The central control (both of the spontaneous
receptor activity and of the input) is, as we shall see, primarily of
a restraining, inhibitory nature. But for the time being let us confine
ourselves to motor organization.
gradients and organelles; it is a structural mosaic equipped with axial
polarity which under normal conditions predetermines the head and tail
region, the blastopores, etc., of the embryo; but it also has striking
regulative properties revealed by experimental manipulation. With
progressive differentiation the regulative properties of the cell
diminish, and its degrees of 'genetic freedom' freeze up. On the level
of the morphogenetic field we again find self-regulating properties --
half the field will still form a complete limb -- and a mosaic-matrix
of cell-populations. The autonomous, self-assertive character of
morphogenetic fields is manifested in grafting and centrifuging
experiments; their dependent, or part character by the trivial fact that
they are kept in their proper size and place in the normally growing
organism. This will sound less of a truism when we turn to the phenomena
of regeneration (Chapter III). We shall find regulative character and
mosaic character, autonomy and subservience, the self-assertion of
the part and its dependence on the whole to be complementary aspects
on every level of the hierarchy in normal development and behaviour;
but also, that under abnormal conditions this ceases to be the case,
and that the part may then assert itself at the expense of the whole,
with sometimes beneficial, mostly destructive, effects.*
NOTES
To
p. 418
. This is, of courre, not meant to
belittle the enormous advantages of sexual over asexual reproduction.
p. 419
. The three-letter 'dictionary', for
instance, is partly a dictionary of synonyms: there are 4^3 = 64 triplets,
but only 20 arnino acids, and many of the latter are represented by more
than one code syllable.
p. 420
. 'Respiration' is an approximate
term. The process is in fact oxidative phosphorylation.
p. 422
. In some, probably extreme cases, the
nuclear changes are even more drastic. The nucleus of the fertilized egg
of the gall-midge contains forty chromosomes, and in the course of the
first few divisions these are faithfully duplicated. But in the fifth
division, only eight sets of chromosomes in the soma cells duplicate in
the orthodox manner; the other thirty-two fail to do so and gradually
dissolve in the cytoplasm. The future germ calls, however, which have
previously been segregated from the rest of the eggs, do not participate
in the fateful fifth division and preserve their chromosome complement
intact. Thus the nuclei of all specialized body cells have only eight
chromosomes, whereas the germ cells have forty. Cf. Fischberg, M. and
Blackler, A. W. (1961).
p. 423
. In a paper read at the British
Association Meeting in August 1962, L. Wolpert suggested that
differentiation resulted from the single cell's tendency to stick on to
that part of the gastrula wall best suited for it (the idea seems to have
been originated by T. Gusthafson). At the same meeting E. N. Willmer showed
that changes in the salt balance of the surrounding medium made amoebae
change from amoeboid to flagellate form, reversibly -- the implication
being that chemical gradients played an important part in the early
stages of differentiation (
New Scientist
, No 303, 6.9.1962, p. 492).
p. 425
. During maturation in the higher species,
the two types of control overlap; and pre-set biological time-clocks
seem to exercise some influence throughout adult life.
p. 428
. In this necessarily simplified
discussion of morphogenetic processes I have made no mention of
cytoplasmic inheritance and other complicating factors, which do not
affect the basic argument of this book.
embryos is G. E. Coghill's monumental study of
ambystoma
, a larval
form of salamander). [1] Coghill published his results in a series
of papers spread over a period of twenty-five years, 1914-39. Since
they are surprisingly seldom quoted outside the technical literature --
presumably because they ran against the behaviourist Zeitgeist -- I must
briefly summarize his conclusions.
was that elementary, local reflexes arise first, and are chained together
at a later stage. Thus the segmental reflex arcs of the earthworm would
develop first, as independent units aligned in a series perpendicular
to its axis, and only later on would they become connected, like rings
hanging from a festoon string, by the spinal cord. Coghill's work showed
that the opposite is true. In the salamander, development starts with
the growth of the motor-tracts of the cord axially from head to tail;
then this central bundle sends out collateral branches into the segmental
muscles, coordinating their actions in primitive, unitary patterns; the
sensory neurons become functional only at a later stage, and the local
reflex-arcs come last, as segregations of 'partial patterns' out of the
'total pattern' which preceded them. The whole development is centrifugal:
the stem precedes the branches, spontaneous undifferentiated movements
involving the whole neuro-muscular apparatus precede differentiated
movement, total responses precede specialized local responses. To give
an example: when the limbs develop, their first movements are entirely
dependent on and synchronized with the movements of the trunk. Only
later on do the limbs begin to move independently; the same applies to
the motions of head, mouth, etc. The growth of the nervous system from
beginning to end is dominated by 'a totally integrated matrix, and not
a progressive integration of primarily individuated units'. The organism
is not a sum of its reflexes, but on the contrary 'the mechanism of the
total pattern is an essential component of the performance of the part,
i.e. the reflex'. The stimulus-response scheme cannot explain even
embryonic behaviour, because movements appear long before the motor
neurons of the reflex arc are connected with the sensory neurons. This
centrifugal mode of development means that the individual acts on its
environment before it reacts to its environment.
development of the individual has been carried, structural provision
has been found for the perpetuation of spontaneity, autonomy, or
initiative as a factor in its behaviour. Any theory of motivation,
therefore, that attributes this function wholly to the environment, is
grossly inadequate.' The idea that instincts are chained reflexes must
be abandoned; instincts represent 'total action patterns in response to
relatively general situations'. Comparing the embryonic development of
ambystoma with that of the human foetus, Coghill sums up:
In conclusion I am convinced by a study of all available records ofWe thus find in the development of the nervous system the same
movement in human foetuses during the first six months, that behaviour
develops in man as it does in ambystoma by the expansion of a total
pattern that is integrated as a whole from the beginning, and by
individuation of partial patterns (reflexes) within the unitary whole.*
principles at work which we have discussed before. The neural plate
starts as a primordium with multipotential cell-populations which
differentiate in a series of steps into the brain, the spinal cord,
and its substructures. The 'wiring diagram' of the organism has a
standardized pattern -- an invariant code; but transplant experiments
again show the great flexibility of the 'neurogenetic skill' which
realizes that pattern. If a limb-bud from a salamander embryo is
transplanted to another embryo's flank, outgrowing nerve-fibres locate
the bud and establish a normal nerve pattern. The bulb-shaped tips of the
outgrowing nerves are apparently guided by submicroscopic structures in
the cell-matrix of the growing bud -- at least according to the current
'contact guidance' theory.
I have called differentiation of structure and integration of function
complementary aspects of a unitary process. But the 'functions' of the
growing embryo are different from the 'functions' of the adult. It has
been shown that the limb-buds and wing buds of chick embryos develop
into almost normal legs and wings if nerves are prevented from entering
them. This does not mean, of course, that differentiation of structure
comes first, and integration of function later on, as a separate act. For
the function of the leg-bud is to grow -- not to walk. Growth is a
function controlled by the genetic code; when growth is completed and
the time has come to walk, the nervous system takes over control; and
if it fails to do so, the muscle tissues will degenerate, as denervated
adult muscles do.*
Locomotor Hierarchies
'Whatever the nature of organizing relations may be,' J. Needham wrote
in 1932, 'they form the central problem of biology, and biology will
be fruitful in the future only if this is recognized. The hierarchy
of relations, from the molecular structure of carbon compounds to the
equilibrium of species and ecological wholes, will perhaps be the leading
idea of the future.' [2]
This prophecy has not come true. The Gestalt school's over-emphasis on
'wholeness', and the behaviourists' over-emphasis on 'simple elementary
processes' -- the so-called S.-R. (stimulus-response) scheme -- created
a controversy based on a fallacious alternative, and prevented a true
appreciation of the multi-layered hierarchic order to be found in all
manifestations of life.
Yet the idea is of course by no means new; hierarchies in nervous function
were proposed by Herbert Spencer in the 1870s, and elaborated by Hughlings
Jackson, Sherrington, and others. [3] The hierarchical character of
skills was demonstrated in great detail by Bryan and Harter in their
study of telegraphy and in Book's study of touch-typing (see below,
pp. 544
ff.) at the turn of the century,
but neither S-R psychologists nor Gestaltists paid attention to
them. Woodger (1929) attempted a formalization, by means of symbolic
logic, of certain types of hierarchies ('divisional hierarchies', 'spatial
hierarchies', 'genetic hierarchies', etc.) which are of somewhat abstract
interest. Heidenhain (1923) [4] proposed a hierarchy of 'histo-systems'
which are 'encapsulated' into one another (e.g. neuro-fibriles, neurons,
nerve fibres). Bertalanffy (1952) tried to make a distinction between
'hierarchies of parts', 'hierarchies of processes', 'hierarchies of
centralization', etc. Tinbergen defined instinct as a hierarchically
organized nervous mechanism -- but his mechanism is fixed and rigid
(see below p. 478). A stimulating discussion of the subject can be found
in Miller, Galanter, and Pribram's remarkable essay on 'Plans and the
Structure of Behaviour' (1960).
'habit family hierarchy', for instance, means just that (the ordering
of a group of interchangeable responses according to their strength),
and is not a hierarchy at all in the sense in which the term is used in
this book. I have summarized what I meant by it in the chapter 'Partness
and Wholeness' (Book One, Chapter XIII). A hierarchy, in this sense, is
not like a row of organ pipes; it is like a tree, arborizing downward. The
structural or functional entities on each level are autonomous sub-wholes
of complex pattern, but are represented on the next higher level as
units. In every organic hierarchy, to paraphrase Gertrude Stein's
statement about the rose, 'a part is a whole is a part is a whole'.
hierarchic order was given by Paul Weiss -- whose experimental work
was a major contribution towards providing the concept with a firm
empirical basis. The quotation which follows is from the celebrated Hixon
Symposium; its vivickss is enhanced by the fact that it is taken from an
ex tempore contribution by Weiss to the discussion of Lashley's paper on
'The Problem of Serial Order in Behaviour'* (my italics):
While the physiologist and psychologist deal with the ready-madeLet me enlarge on some of these points and add a few facts which have
machine of the nervous system and can add to it as many properties as
he thinks necessary, the embryologist must explain just how such an
immensely intricate, yet orderly, thing can develop. These studies
are still in their infancy, but a few things have already come out
. . . for instance, the relative autonomy of structured patterns
of activity, and the hierarchical principle of their organization
. . . . The nervous system is not one big monotonic pool whose elements
can be freely recombined in any number of groupings, thereby giving an
infinite variety of nervous responses. This used to be the old idea of
the associationists, and it is utterly incompatible with what we have
learned about the development of the nervous system and its function
in animals.
The working of the central nervous system is a hierarchic affair in
which functions at the higher levels do not deal directly with the
ultimate structural units, such as neurons or motor units, but
operate by activating lower patterns that have their own relatively
autonomous structural unity. The same is true for the sensory input
which . . . operates by affecting, distorting, and somehow modifying the
pre-existing, preformed patterns of central co-ordination. . . . The
final output is then the outcome of this hierarchical passing down
of distortions and modifications of intrinsically preformed patterns
of excitation, which are in no way replicas of the input. The
structure of the input does not produce the structure of the output,
but merely modifies intrinsic nervous activities that have a structural
organization of their own. This has been proved by observation and
experiment. Coghill has shown that the motor patterns of the animal
develop prior to the development of sensory innervation. I have shown,
as others have, that the removal of the sensory innervation does
not abolish the co-ordination of motor activities. Moreover,
coordinated motor functions of limbs and other parts develop even
if these parts have been experimentally prevented from ever becoming
innervated by sensory fibres. Therefore, the sensory pathway can have
nothing to do with the structure of the motor response. There are
still some authors who try to save the old associationist idea that
actually the input shapes the structure of the output. I think that
they are fighting a losing fight, and I think that today's discussion
ought to have given them the coup de grâce.
Intrinsic automatic rhythms have been shown, for instance, by Adrian
in the brain stem of the goldfish and in insect ganglia, by Prosser
in other arthropods, by Bremer and by von Holst in the spinal cord,
and by Bethe in jellyfish. I have shown experimentally that any group
of bulbar or spinal nerve cells taken from vertebrates, if deprived of
their structural bonds of restraining influences and allowed to undergo
a certain degree of degradation, will display permanent automatic,
rhythmic, synchronized activity of remarkable regularity. Rhythmic
activity, therefore, seems a basic property of pools of nervous
elements . . . . The rhythm is not something generated through an
input rhythm; but is itself a primary rhythm which may be released
and even speeded up or retarded by the input, but is not derived
from the input. So we have experimental evidence that rhythmic
automatism, autonomy of pattern, and hierarchical organization are
primary attributes of even the simplest nervous systems , and I
think that this unifies our view of the nervous system. [5]
emerged since.
In the first place it has been found that intrinsic, rhythmic activity
of an autonomous character is not confined to motor nerves, but
that 'receptors also are spontaneously active even in the absence of
stimulation from environment.' [6] This spontaneous receptor activity,
while modified by environmental events, is under efferent control from
the central nervous system. The central control (both of the spontaneous
receptor activity and of the input) is, as we shall see, primarily of
a restraining, inhibitory nature. But for the time being let us confine
ourselves to motor organization.
Other books
Son of Sun (Forgotten Gods (Book 2)) by Clair, Rosemary
Bound, Spanked and Loved: Fourteen Kinky Valentine's Day Stories by Sierra Cartwright, Annabel Joseph, Cari Silverwood, Natasha Knight, Sue Lyndon, Emily Tilton, Cara Bristol, Renee Rose, Alta Hensley, Trent Evans, Ashe Barker, Katherine Deane, Korey Mae Johnson, Kallista Dane
The Sword That Cut the Burning Grass by Dorothy Hoobler
An Honest Deception by Alicia Quigley
Captivated by Susan Scott Shelley
Caught in a Bind by Gayle Roper
Night Sky (Satan's Sinners MC Book 3) by Kay, Colbie
The Bretwalda (The Casere Book 4) by Michael O'Neill
The Dying Hours by Mark Billingham
The Devil's Alphabet by Daryl Gregory