In The Blink Of An Eye (36 page)
Read In The Blink Of An Eye Online
Authors: Andrew Parker

At the beginning of this chapter we examined light perceivers that could detect light levels but not form visual images. They were not eyes. But some light perceivers were more efficient than others, and it is likely that a more efficient type originated from a less efficient type
during its evolution. To make their predictions, Nilsson and Pelger applied this logic.
during its evolution. To make their predictions, Nilsson and Pelger applied this logic.
A patch of light-sensitive skin was used as a starting point. This dents inwards, and becomes increasingly infolded to form detectors escalating in their sensitivity to the direction of light. This assumption is quite acceptable since all the intermediate stages can be found functioning in animals today. It is important that each link in the chain
can
exist in its own right. The opposite of this was once used to criticise evolution, and even clouded the thoughts of Darwin himself, as suggested in the epigraph at the beginning of this chapter. To justify this further, we can explain why all animals don't possess the theoretical ultimate eye. The intermediate stages, or conceptually substandard visual organs,
do
exist today because their host animals cannot handle the information loads supplied by the next conceivable stage on the road to a fully formed camera-type eye - Darwin had no reason to be concerned. Back on the evolutionary road, we have reached a âcup eye' that cannot form proper images. We have also reached a junction. Close the entrance to the cup even more and we have the pinhole eye of nautilus. Then again, begin to grow a lens and another path has been taken - the path to the camera-type eye typical of vertebrates.
can
exist in its own right. The opposite of this was once used to criticise evolution, and even clouded the thoughts of Darwin himself, as suggested in the epigraph at the beginning of this chapter. To justify this further, we can explain why all animals don't possess the theoretical ultimate eye. The intermediate stages, or conceptually substandard visual organs,
do
exist today because their host animals cannot handle the information loads supplied by the next conceivable stage on the road to a fully formed camera-type eye - Darwin had no reason to be concerned. Back on the evolutionary road, we have reached a âcup eye' that cannot form proper images. We have also reached a junction. Close the entrance to the cup even more and we have the pinhole eye of nautilus. Then again, begin to grow a lens and another path has been taken - the path to the camera-type eye typical of vertebrates.
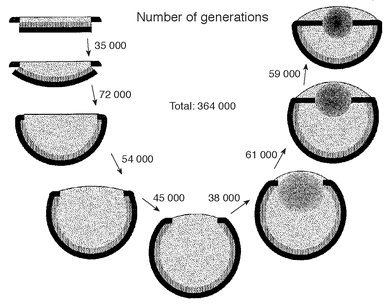
Figure 7.16
Nilsson and Pelger's predicted evolution of a camera-type eye, like that of a fish. The sequence begins with a flat patch of light-sensitive cells sandwiched between a transparent protective layer and a layer of dark pigment. A graded-index lens appears at stage 6. Reproduced from a 1994 paper by Nilsson and Pelger with permission from the authors.
Nilsson and Pelger's predicted evolution of a camera-type eye, like that of a fish. The sequence begins with a flat patch of light-sensitive cells sandwiched between a transparent protective layer and a layer of dark pigment. A graded-index lens appears at stage 6. Reproduced from a 1994 paper by Nilsson and Pelger with permission from the authors.
Nilsson and Pelger were more than realistic in assuming that a light receptor will change by just 1 per cent of its length, width or protein density during each evolutionary step in the eye direction. But even with such a pessimistic approach, the whole sequence from light-sensitive patch to the eye of a fish would require only two thousand of these tiny modifications in sequence. That may not seem enough, but as Michael Land and Dan-Eric Nilsson point out, if two thousand sequential modifications of 1 per cent are applied to the length of a finger, then it becomes long enough to bridge the Atlantic Ocean.
We know that proteins need not evolve from their chemical beginnings. A study of flatworms revealed that similar proteins exist in the eyespots (not true eyes)
and
touch/chemical detectors. In the eyespots, these are the proteins that react to light, comparable to those in the retina of an eye. So a head start may be gained towards eye evolution by borrowing the proteins of other detectors.
and
touch/chemical detectors. In the eyespots, these are the proteins that react to light, comparable to those in the retina of an eye. So a head start may be gained towards eye evolution by borrowing the proteins of other detectors.
Now for the calculation of time needed for these modifications to take place, which is really what we are interested in. Again, caution was the name of the game when Nilsson and Pelger made their assumption about the slowest rate of evolution - a 0.005 per cent modification from one generation to the next. In reality, the rate would probably be faster. For instance, the light receptor pigments of modern crustaceans show an evolution that is considerably more
rapid
than expected. And verily the word âpessimistic' entered the title of Nilsson and Pelger's original paper, which made their result seem even more remarkable. They found that the eye of a fish could evolve from its rudimentary
beginnings in less than 400,000 generations. Assuming each generation is completed within a year, this result suggests that an efficient, image-forming eye can evolve in less than half a million years. Now that really is a blink of an eye on the geological timescale.
rapid
than expected. And verily the word âpessimistic' entered the title of Nilsson and Pelger's original paper, which made their result seem even more remarkable. They found that the eye of a fish could evolve from its rudimentary
beginnings in less than 400,000 generations. Assuming each generation is completed within a year, this result suggests that an efficient, image-forming eye can evolve in less than half a million years. Now that really is a blink of an eye on the geological timescale.
This is a camera-type eye and we have established that the first eye was compound. But in their definitive book on the optics of animal eyes, called
Animal Eyes
, Michael Land and Dan-Eric Nilsson were beginning to picture the evolutionary sequence of the compound eye. They claimed that arthropods âprobably originated from a worm-like ancestor that already possessed a rudimentary compound eye - possibly a loose collection of eyespots'. Independently, the Australian biologist Richard Smith mapped the changes needed to form the compound âeye' of a bristle worm. A loose collection of eyespots also appeared in Smith's sequence. And the number of links expected in the chain leading to a fully functioning eye was on a par with those in Nilsson and Pelger's predictions for the camera-type eye.
Animal Eyes
, Michael Land and Dan-Eric Nilsson were beginning to picture the evolutionary sequence of the compound eye. They claimed that arthropods âprobably originated from a worm-like ancestor that already possessed a rudimentary compound eye - possibly a loose collection of eyespots'. Independently, the Australian biologist Richard Smith mapped the changes needed to form the compound âeye' of a bristle worm. A loose collection of eyespots also appeared in Smith's sequence. And the number of links expected in the chain leading to a fully functioning eye was on a par with those in Nilsson and Pelger's predictions for the camera-type eye.
Like the proteins of the retina, other parts of the body involved in the process of light perception seem quite accommodating to these calculations on the eyes themselves. Nilsson and Pelger's time prediction would be meaningless if development of the visual processing centre in the brain was lagging behind that of the eye. In 1959 the biologist von Bekesy demonstrated that the effects caused by sound can be mimicked by vibrating the skin. This demonstrated that the ear and skin shared certain common features, namely nerves, in the processing of sensory information. But what does this mean for the evolution of the eye? Well, it is conceivable that nerves used by one sense can be âupgraded' for use by two senses. And if the senses of hearing and touch can share features, then so might vision and touch. In this way, the nerves needed to service an eye would not have to evolve from a vestigial beginning - they would have a head start. Then there is a possible helping hand in the brain department. Parts of the brain, it appears, may be capable of converting from touch to vision. Dan-Eric Nilsson suggested that the compound âeye' of ark clams and bristle worms evolved from chemical detectors that were inhibited by light. So the evolution of the
eye itself
appears to be the limiting factor, or at the back of the pack, on the evolutionary road to vision - the remainder of
the system can simply be adopted. Indeed, there were other sense organs surrounding the eyes of trilobites, and the original light perceivers may have borrowed nerves from these.
eye itself
appears to be the limiting factor, or at the back of the pack, on the evolutionary road to vision - the remainder of
the system can simply be adopted. Indeed, there were other sense organs surrounding the eyes of trilobites, and the original light perceivers may have borrowed nerves from these.
Now we can calm our own nerves that may have been jangling while we gave the compound eye just one million years to evolve - at least if it was to fit with our fossil evidence. It seems that our demand has been met - one million years is plenty of time for an eye to evolve. Now we can paint a picture of 544 million years ago, where light sensitive patches were evident in the ancestors of the Cambrian trilobites. Then we can paint another picture of 543 million years ago, just the other side of the Cambrian border, where a trilobite proudly flaunts its eyes. Between the two pictures the light-sensitive patch had evolved into an eye.
Between 544 and 543 million years ago a revolution took place. During this one million year period, vision was born.
We are now in a position to interpret the statement âHow ancient already in the Lower Cambrian must the compound eye have been' made by Frank Raw. Yes, the compound eye and vision were well developed in the Lower Cambrian. But no, it was not ancient - it was contemporary. And it became the new fashion.
There was always going to be one moment in history when the eye suddenly appeared on Earth, as if out of nowhere. Now we can identify that moment. And a really important point to bear in mind at all times is that light-sensitive patches and other stages of rudimentary light receptors are not eyes. While only these patches existed, when eyes were awaiting their introduction to Earth, there was no such thing as vision.
We now know that eyes existed at the very beginning of the Cambrian . . . but not before. These two facts could be as important as each other. Considered together, they describe the introduction of a sense. Not just any old sense, but the most powerful sense or stimulus to animal behaviour and evolution in sunlit environments. And a sunlit environment is where the Burgess Shale and other well-known Cambrian animals lived. It also played host to the Cambrian explosion.
Extrapolating further, there are lifestyles that can be reconstructed based on the optics of eyes. The architecture of eyes alone can provide
information on how animals lived. For instance, the position of the eyes in the head can reveal the position of the animal in the food chain. Eyes positioned at the sides of the head, facing sideways like those of a rabbit, can scan a wide angle and spot movement from nearly all directions. The movement pursued in this case is that of predators - eyes of this type belong mainly to plant eaters. In contrast, eyes positioned together at the front of the head, facing forward like those of an owl, see less of the environment but are better for pinpointing targets and judging the distance between them. These eyes generally belong to meat eaters. But this is a theme for another chapter.
information on how animals lived. For instance, the position of the eyes in the head can reveal the position of the animal in the food chain. Eyes positioned at the sides of the head, facing sideways like those of a rabbit, can scan a wide angle and spot movement from nearly all directions. The movement pursued in this case is that of predators - eyes of this type belong mainly to plant eaters. In contrast, eyes positioned together at the front of the head, facing forward like those of an owl, see less of the environment but are better for pinpointing targets and judging the distance between them. These eyes generally belong to meat eaters. But this is a theme for another chapter.
8
The Killer Instinct
A little alarm now and then keeps life from stagnation
F. BURNEY (Mme d'Arbley),
Camilla
(1796)
Camilla
(1796)
Â
Â
Â
THE LAWS OF LIFE
For the survival of animals everywhere
Â
CONTENTS
Â
Basic Rules
1.
Every man for himself: stay alive!
Every man for himself: stay alive!
1a. Avoid being eaten
1b. âEat'
2.
For the good of one's kind.
For the good of one's kind.
2a. Breed
2b. Find a niche and protect it
2c. Adapt to changes in the environment
Lifestyle
1. Predator
2. Prey
Tactics
1. Conspicuousness
2. Crypsis/illusiveness
3. Genuine strength/ability
The previous chapter could be viewed as âend of story'. Certainly, there is considerable evidence within that chapter suitable for the Cambrian files. But it is too early to jump to conclusions just yet, for there is something else to consider, a subject that has raised its head, either plainly or rather more cryptically, in every chapter so far. In each case it merged into the background as quickly as it appeared. Before ending our Cambrian investigation, we should introduce
predators
into the evidence.
predators
into the evidence.
The first rule of animal survival is to stay alive. The other rules, such as feeding and breeding, are academic if this first rule is not followed. But from the beginning we must distinguish between an individual and a species. A species is a collection of like individuals, which interbreed in their natural environment. Staying alive and feeding are factors that directly affect individuals, then indirectly species. Breeding and niche occupations are concerns for the long-term survival of the species. Of course animals don't really receive rules - in reality the rules for their survival are the selective pressures for evolution, invisible forces acting on the genes, carrying messages for enhanced survival. And selective pressures act directly upon individuals, not species, so even the species-level survival factors are relayed through individuals.
The first basic rule of species survival - for individuals to stay alive - will form the subject of this chapter. And more specifically, I will centre on the most important aspect of that rule, to avoid being eaten. This chapter is a stage for the predators. And, in keeping with the previous chapters, the stage will have a space
and
a time dimension.
and
a time dimension.
Before launching into the world of
T. rex
and the like, I will make a brief disclaimer relating to The Laws of Life outlined on the previous page. These are the general rules but do not cover all possibilities, particularly those less common natural catastrophes. Some things are beyond evolution, such as meteor impacts, sudden ice ages, and disease. Disease is density dependent, and so it is a factor operating at the species level. On the one hand, species can become too successful for their own good. From another viewpoint, this is just evolution maintaining biodiversity, preventing one species from taking over the world. But in general, biodiversity is maintained by
all
branches of the evolutionary tree adhering to The Laws of Life. A predator does not become an overnight success by growing bigger teeth. The other side of the coin
is the âAvoid being eaten' alarm for its prey species, which favours genetic mutations for stronger armour. Cichlid fishes feed on snails, and where the fishes evolve stronger teeth, the snails simply evolve harder shells. Evolution can take animals down different roads. There are roads to predation and there are roads to prey, with the predator and prey roads running between. But all roads are endless, and animals are continuously moving along
all
of them. However, all animals today are travelling along an established evolutionary road - snails already possess armour that may yet become reinforced.
T. rex
and the like, I will make a brief disclaimer relating to The Laws of Life outlined on the previous page. These are the general rules but do not cover all possibilities, particularly those less common natural catastrophes. Some things are beyond evolution, such as meteor impacts, sudden ice ages, and disease. Disease is density dependent, and so it is a factor operating at the species level. On the one hand, species can become too successful for their own good. From another viewpoint, this is just evolution maintaining biodiversity, preventing one species from taking over the world. But in general, biodiversity is maintained by
all
branches of the evolutionary tree adhering to The Laws of Life. A predator does not become an overnight success by growing bigger teeth. The other side of the coin
is the âAvoid being eaten' alarm for its prey species, which favours genetic mutations for stronger armour. Cichlid fishes feed on snails, and where the fishes evolve stronger teeth, the snails simply evolve harder shells. Evolution can take animals down different roads. There are roads to predation and there are roads to prey, with the predator and prey roads running between. But all roads are endless, and animals are continuously moving along
all
of them. However, all animals today are travelling along an established evolutionary road - snails already possess armour that may yet become reinforced.
Central to this book so far have been the subjects of light and vision. When superimposed on to The Laws of Life, their capacity will become evident. Specifically, they fall into the âTactics' section. Consider the Hawaiian unicorn fish with its conspicuous yellow spine near its tail. The spine serves to protect the fish from predators and competitors, and consequently the unicorn fish avoids being eaten and protects its niche. But the unicorn fish rarely calls upon its spine because in reality this armament is only an ornament. Here the messenger is light. Potential predators and competitors see the armoury and have second thoughts.
Other books
Death of a Commuter by Bruce, Leo
Primal Passion by Mari Carr
Supernatural: Carved in Flesh by Waggoner, Tim
Ripe: A Stepbrother Pregnancy Romance by Thorn, Kate
Creators by Paul M. Johnson
Lilies and Lies by Mary Manners
Dance of Demons by Gary Gygax
Dr. Futurity (1960) by Philip K Dick
The Seduction - Art Bourgeau by Art Bourgeau
Addison Cooke and the Treasure of the Incas by Jonathan W. Stokes