Bully for Brontosaurus (18 page)
Read Bully for Brontosaurus Online
Authors: Stephen Jay Gould

But what is so wrong with these evolutionary ladders? Surely we can trace an unbroken continuity from
Hyracotherium
to modern horses. Yes, but continuity comes in many more potential modes than the lock step of the ladder. Evolutionary genealogies are copiously branching bushes—and the history of horses is more lush and labyrinthine than most. To be sure,
Hyracotherium
is the base of the trunk (as now known), and
Equus
is the surviving twig. We can, therefore, draw a pathway of connection from a common beginning to a lone result. But the lineage of modern horses is a twisted and tortuous excursion from one branch to another, a path more devious than the road marked by Ariadne’s thread from the Minotaur at the center to the edge of our culture’s most famous labyrinth. Most important, the path proceeds not by continuous transformation but by lateral stepping (with geological suddenness when punctuated equilibrium applies, as in this lineage, at least as read by yours truly, who must confess his bias as coauthor of the theory).
Each lateral step to a new species follows one path among several alternatives. Each extended lineage becomes a set of decisions at branching points—only one among hundreds of potential routes through the labyrinth of the bush. There is no central direction, no preferred exit to this maze—just a series of indirect pathways to every twig that ever graced the periphery of the bush.
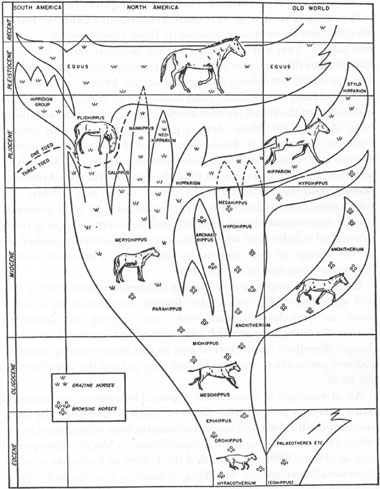
The evolution of horses depicted as at least a modest bush by G.G. Simpson in 1951.
NEG. NO. 328907. COURTESY DEPARTMENT OF LIBRARY SERVICES, AMERICAN MUSEUM OF NATURAL HISTORY
.
As an example of distortions imposed by converting tortuous paths through bushes into directed ladders, consider the men associated with the two classical iconographies reproduced here. When Huxley made his formal capitulation to Marsh’s interpretation in print (1880), he extended the ladder of horses as a metaphor for all vertebrates. Speaking of modern reptiles and teleost fishes, Huxley wrote (1880, p. 661): “They appear They appear to me to be off the main line of evolution—to represent, as it were, side tracks starting from certain points of that line.” But teleosts (modern bony fishes) are an enormously successful group. They stock the world’s oceans, lakes, and rivers and maintain nearly 100 times as many species as primates (and more than all mammals combined). How can we call them “off the main line” just because we can trace our own pathway back to a common ancestry with theirs more than 300 million years ago?
W. D. Matthew slipped into an equally biased assessment of value because his designation of one pathway as a ladder forced an interpretation of all others as diversions. Matthew (1926, p. 164) designated his ladder as the “direct line of succession,” but acknowledged that “there are also a number of side branches, more or less closely related.” Three pages later, Matthew adds the opprobrium of near indecency to his previous charge of mere laterality, as he describes (p. 167) “a number of side branches leading up in a similar manner to aberrant specialized Equidae now extinct.” But in what way are extinct lineages more specialized than a modern horse or in any sense more peculiar? Their historical death is the only possible rationale for a designation of aberrancy, but more than 99 percent of all species that ever lived are extinct—and disappearance cannot be the biological equivalent of a scarlet letter. We might as well call modern horses aberrant because, much to Montezuma’s later sorrow, they became extinct in the land of their birth.
Yet we have recognized the bushiness of horse evolution from the very beginning. How else did Marsh forestall Huxley but by convincing him that his European “genealogy” of horses formed a stratigraphic sequence of discontinuous stages, falsely linking several side branches that had disappeared without issue?
As an example of bushiness, and a plug for the value of appropriate metaphors in general, consider the finest book on the evolution of horses ever written for popular audiences—G. G. Simpson’s
Horses
(1951). Simpson redrew the genealogy of horses as a modest bush with no preferred main line. He also criticized the conceptual lock imposed by the bias of the ladder when he noted that modern one-toed horses are a side branch and extinct three-toed creatures the main line (if any center can be designated at all).
As nearly as there is a straight line in horse evolution, it culminated and ended with these animals [the three-toed anchitheres], which, like their ancestors, were multiple-toed browsers. From this point of view, it is the line leading to modern horses that was the side branch, even though it outlasted the straighter line of horse evolution [p. 130].
Yet Simpson, who held a lifelong commitment to the predominant role of evolution by transformational change within populations rather than by accumulation across numerous events of discrete, branching speciation, could not entirely let go of biases imposed by the metaphor of the ladder. In one revealing passage, he accepts bushiness, but bemoans the complexities thus introduced, as though they clouded evolution’s essence of transformational change:
Miohippus
…intergraded with several different descendant groups. It is sad that this introduces possible confusion into the story, but there is not much point in criticizing nature for something that happened some millions of years ago. It would also be foolish to try to ignore the complications, which did occur and which are a very important part of the record.
But these “complications” are not a veil upon the essence of lineal descent; they are the primary stuff of evolution itself.
Moreover, Simpson restricted his bushiness as much as possible and retained linearity wherever he could avoid an inference of branching. In particular, he proposes the specific and testable hypothesis (see his illustration) that the early part of the record—the sequence of
Hyracothenum—Orohippus—Epihippus—Mesohippus—Miohippus—Hypohippus
—tells a story of linear descent, only later interrupted by copious branching among three-toed browsers: “The line from
Eohippus
to
Hypohippus
, for example, exemplifies a fairly continuous phyletic evolution” (p. 217). Simpson especially emphasizes the supposedly gradual and continuous transformation from
Mesohippus
to
Miohippus
near the top of this sequence:
The more progressive horses of the middle Oligocene and all the horses of the late Oligocene are placed by convention in a separate genus,
Miohippus
. In fact
Mesohippus
and
Miohippus
intergrade so perfectly and the differences between them are so slight and variable that even experts find it difficult, at times nearly impossible, to distinguish them clearly.
The enormous expansion of collections since Simpson proposed this hypothesis has permitted a test by vertebrate paleontologists Don Prothero and Neil Shubin. Their results falsify Simpson’s gradual and linear sequence for the early stages of horse evolution and introduce extensive bushiness into this last stronghold of the ladder.
Prothero and Shubin have made four major discoveries in the crucial segment of history that Simpson designated as the strongest case for a gradualistic sequence of lineal transformation—the transition from
Mesohippus
to
Miohippus
.
1. Previous experts were so convinced about the imperceptibly gradual transition between these two genera that they declared any search for distinguishing characters as vain, and arbitrarily drew the division between
Mesohippus
and
Miohippus
at a stratigraphic boundary. But far richer material available to Prothero and Shubin has permitted the identification of characters that cleanly distinguish the two genera. (Teeth are the hardest part of a vertebrate skeleton and the fossil record of mammals often contains little else. A technical course in the evolution of mammals is largely an exercise in the identification of teeth, and an old professional quip holds that mammalian evolution is the interbreeding of two sets of teeth to produce some slightly modified descendant choppers.
Miohippus
and
Mesohippus
do not have distinctive dentitions, and previous failure to find a clear separation should not surprise us. The new material is rich in skull and limb bones.) In particular, Prothero and Shubin found that
Miohippus
develops a distinctive articulation, absent in ancestral
Mesohippus
, between the enlarging third metatarsal (the foot bone of the digit that will become the entire hoof of modern horses) and the cuboid bone of the tarsus (ankle) above.
2.
Mesohippus
does not turn into
Miohippus
by insensible degrees of gradual transition. Rather,
Miohippus
arises by branching from a
Mesohippus
stock that continues to survive long afterward. The two genera overlap in time by at least 4 million years.
3. Each genus is itself a bush of several related species, not a rung on a ladder of progress. These species often lived and interacted in the same area at the same time (as different species of zebra do in Africa today). One set of strata in Wyoming, for example, has yielded three species of
Mesohippus
and two of
Miohippus
, all contemporaries.
4. The species of these bushes tend to arise with geological suddenness, and then to persist with little change for long periods. Evolutionary change occurs at the branch points themselves, and trends are not continuous marches up ladders, but concatenations of increments achieved at nodes of branching on evolutionary bushes. Of this phenomenon Prothero and Shubin write:
There is no evidence of long-term changes within these well-defined species [of
Mesohippus
and
Miohippus
] through time. Instead, they are strikingly static through millions of years. Such stasis is apparent in most Neogene [later] horses as well, and in
Hyracotherium
. This is contrary to the widely-held myth about horse species as gradualistically-varying parts of a continuum, with no real distinctions between species. Throughout the history of horses, the species are well-marked and static over millions of years. At high resolution, the gradualistic picture of horse evolution becomes a complex bush of overlapping, closely related species.
Bushiness now pervades the entire phylogeny of horses.
We can appreciate this fundamental shift in iconography and meaning, but where is the “precious irony” that I promised? What is “life’s little joke” of my title? Simply this. The model of the ladder is much more than merely wrong. It never could provide the promised illustration of evolution progressive and triumphant—for
it could only be applied to unsuccessful lineages
.
Bushes represent the proper topology of evolution. Ladders are false abstractions, made by running a steamroller over a labyrinthine pathway that hops from branch to branch through a phylogenetic bush, We cannot force a successful bush of evolution into a ladder because we may follow a thousand pathways through the maze of twigs, and we cannot find a criterion for preferring one route over another. Who ever heard of the evolutionary trend of rodents or of bats or of antelopes? Yet these are the greatest success stones in the history of mammals. Our proudest cases do not become our classic illustrations because we can draw no ladder of progress through a vigorous bush with hundreds of surviving twigs.
But consider the poor horses. Theirs was once a luxuriant bush, yet they barely survive today. Only one twig (the genus
Equus
, with horses, zebras, and asses) now carries all the heritage of a group that once dominated the history of hoofed mammals—and with fragility at that, for
Equus
died in the land of its birth and had to be salvaged from a stock that had migrated elsewhere. (In a larger sense, horses form one of three dwindling lines—tapirs and rhinos are the others—that now represent all the diversity of the formerly dominant order Perissodactyla, or odd-toed ungulates, among hoofed mammals. This mighty group once included the giant titanotheres, the clawed chalicotheres, and
Baluchitherium
, the largest land mammal that ever lived. It now hangs on as a remnant in a world increasingly dominated by the Artiodactyla, or even-toed ungulates—cows, deer, antelope, camels, hippos, giraffes, pigs, and their relatives.)
This is life’s little joke. By imposing the model of the ladder upon the reality of bushes, we have guaranteed that our classic examples of evolutionary progress can only apply to unsuccessful lineages on the very brink of extermination—for we can linearize a bush only if it maintains but one surviving twig that we can falsely place at the summit of a ladder. I need hardly remind everybody that at least one other mammalian lineage, preeminent among all in our attention and concern, shares with horses the sorry state of reduction from a formerly luxuriant bush to a single surviving twig—the very property of extreme tenuousness that permits us to build a ladder reaching only to the heart of our own folly and hubris.