Why Evolution Is True (18 page)
Read Why Evolution Is True Online
Authors: Jerry A. Coyne

There’s a poignant footnote to this story. When Robert Scott’s party was found in 1912, frozen to death after their unsuccessful attempt to be the first at the South Pole (the Norwegian Roald Amundsen got there a bit earlier), thirty-five pounds of
Glossopteris
fossils lay next to their bodies. Despite having discarded much of their equipment in a desperate attempt to stay alive, the party physically dragged these heavy rocks on hand sledges, doubtlessly realizing their scientific value. They were the first specimens of
Glossopteris
found in Antarctica.
Glossopteris
fossils lay next to their bodies. Despite having discarded much of their equipment in a desperate attempt to stay alive, the party physically dragged these heavy rocks on hand sledges, doubtlessly realizing their scientific value. They were the first specimens of
Glossopteris
found in Antarctica.
The evidence for evolution from patterns of life on continents is strong, but that from life on islands is, as we shall see, even stronger.
IslandsREALIZING THAT THE DISTRIBUTION of species on islands provides conclusive proof of evolution was one of the greatest pieces of sleuthing in the history of biology. This too was the work of Darwin, whose ideas still loom mightily over the field of biogeography. In chapter 12 of
The Origin,
Darwin reports fact after fact, painstakingly gathered over years of observation and correspondence, building his case like a brilliant lawyer. When I teach the evidence for evolution to my students, this is my favorite lecture. It’s an hourlong mystery story, an accumulation of seemingly disparate data that in the end resolves into an airtight case for evolution.
The Origin,
Darwin reports fact after fact, painstakingly gathered over years of observation and correspondence, building his case like a brilliant lawyer. When I teach the evidence for evolution to my students, this is my favorite lecture. It’s an hourlong mystery story, an accumulation of seemingly disparate data that in the end resolves into an airtight case for evolution.
But before we get to the evidence, we need to distinguish two types of islands. The first are
continental islands:
those islands once connected to a continent but later separated either by rising sea levels that flooded former land bridges or by moving continental plates. These islands include, among many others, the British Isles, Japan, Sri Lanka, Tasmania, and Madagascar. Some are old (Madagascar parted from Africa about 160 million years ago), others much younger (Great Britain separated from Europe around 300,000 years ago, probably during a catastrophic flood spilling from a large, dammed-up lake to the north).
Oceanic islands,
on the other hand, are those that were never connected to a continent; they arose from the seafloor, initially bereft of life, as growing volcanoes or coral reefs. These include the Hawaiian Islands, the Galapagos archipelago, St. Helena, and the Juan Fernández group described at the beginning of this chapter.
continental islands:
those islands once connected to a continent but later separated either by rising sea levels that flooded former land bridges or by moving continental plates. These islands include, among many others, the British Isles, Japan, Sri Lanka, Tasmania, and Madagascar. Some are old (Madagascar parted from Africa about 160 million years ago), others much younger (Great Britain separated from Europe around 300,000 years ago, probably during a catastrophic flood spilling from a large, dammed-up lake to the north).
Oceanic islands,
on the other hand, are those that were never connected to a continent; they arose from the seafloor, initially bereft of life, as growing volcanoes or coral reefs. These include the Hawaiian Islands, the Galapagos archipelago, St. Helena, and the Juan Fernández group described at the beginning of this chapter.
The “island” argument for evolution starts with the following observation: oceanic islands are missing many types of native species that we see on both continents and continental islands. Take Hawaii, a tropical archipelago whose islands occupy about 6,400 square miles, only slightly smaller than the state of Massachusetts. While the islands are well stocked with native birds, plants, and insects, they completely lack native freshwater fish, amphibians, reptiles, and land mammals. Napoleon’s island of St. Helena and the archipelago of Juan Fernandez lack these same groups, but still have plenty of endemic plants, birds, and insects. The Galapagos Islands do have a few native reptiles (land and marine iguanas, as well as the famous giant tortoises), but they too are missing native mammals, amphibians, and freshwater fish. Over and over again, on the oceanic islands that dot the Pacific, the South Atlantic, and the Indian Ocean, one sees a pattern of missing groups—more to the point, the
same
missing groups.
same
missing groups.
At first blush, these absences seem bizarre. If you look at even a very small patch of a tropical continent or a continental island, say in Peru, New Guinea, or Japan, you’ll find plenty of native fish, amphibians, reptiles, and mammals.
As Darwin noted, this disparity is hard to explain under a creationist scenario: “He who admits the doctrine of creation of each separate species, will have to admit, that a sufficient number of the best adapted plants and animals have not been created on oceanic islands.” But how do we know that mammals, amphibians, freshwater fish, and reptiles really are
suited
to oceanic islands? Maybe the creator didn’t put them there because they wouldn’t do well. One obvious reply is that
continental
islands do have these animals, so why would a creator put different types of animals on continental versus oceanic islands? How the island was formed shouldn’t make a difference. But Darwin ends the sentence given above with an even better response: “... for man has unintentionally stocked them from various sources far more fully and perfectly than nature.”
suited
to oceanic islands? Maybe the creator didn’t put them there because they wouldn’t do well. One obvious reply is that
continental
islands do have these animals, so why would a creator put different types of animals on continental versus oceanic islands? How the island was formed shouldn’t make a difference. But Darwin ends the sentence given above with an even better response: “... for man has unintentionally stocked them from various sources far more fully and perfectly than nature.”
In other words, mammals, amphibians, freshwater fish, and reptiles often do very well when humans introduce them to oceanic islands. In fact, they often take over, wiping out native species. Introduced pigs and goats have overrun Hawaii, making meals of native plants. Introduced rats and mongooses have destroyed or endangered many of Hawaii’s spectacular birds. The cane toad, a huge poisonous amphibian native to tropical America, was introduced to Hawaii in 1932 to control beetles on sugarcane. The toads are now a pest, breeding prolifically and killing cats and dogs who mistake them for a meal. The Galapagos Islands have no native amphibians, but an Ecuadorian tree frog, introduced in 1998, has established itself on three islands. On São Tome, the volcanic island off the west coast of Africa where I collect fruit flies for my own research, black cobras have been introduced—perhaps accidentally—from the African mainland. They have done so well that we simply won’t work in certain areas of the island, as the cobras are so numerous that we can encounter several dozen of these deadly and aggressive snakes in a single day. Land mammals do well on islands too—introduced goats helped Alexander Selkirk stay alive on Más a Tierra, and they also thrive on St. Helena. Throughout the world the story is the same: humans introduce species to oceanic islands where they did not exist, and these species displace or destroy native forms. So much for the argument that oceanic islands are somehow unsuitable for mammals, amphibians, reptiles, and fish.
The next step of the argument is this: although oceanic islands lack many basic kinds of animals, the types that
are
found there are often present in profusion, comprising many similar species. Take the Galapagos. Among its thirteen islands there are twenty-eight species of birds found nowhere else. And of these twenty-eight, fourteen belong to a single group of closely related birds: the famous Galapagos finches. No continent or continental island has a bird fauna so heavily dominated by finches. Yet despite their shared finchlike traits, the Galapagos group is ecologically quite diverse, with different species specializing on foods as different as insects, seeds, and the eggs of other species. The “carpenter finch” is one of those rare species that uses tools—in this case a cactus spine or twig to pry insects from trees. Carpenter finches fill the ecological role of woodpeckers, which don’t live in the Galapagos. And there’s even a “vampire finch” that pecks wounds on the rear ends of seabirds and then laps up the blood.
are
found there are often present in profusion, comprising many similar species. Take the Galapagos. Among its thirteen islands there are twenty-eight species of birds found nowhere else. And of these twenty-eight, fourteen belong to a single group of closely related birds: the famous Galapagos finches. No continent or continental island has a bird fauna so heavily dominated by finches. Yet despite their shared finchlike traits, the Galapagos group is ecologically quite diverse, with different species specializing on foods as different as insects, seeds, and the eggs of other species. The “carpenter finch” is one of those rare species that uses tools—in this case a cactus spine or twig to pry insects from trees. Carpenter finches fill the ecological role of woodpeckers, which don’t live in the Galapagos. And there’s even a “vampire finch” that pecks wounds on the rear ends of seabirds and then laps up the blood.
Hawaii has an even more spectacular radiation of birds, the honeycreepers. When the Polynesians arrived in Hawaii about fifteen hundred years ago, they found about 140 species of native birds (we know this from studies of bird “subfossils”: bones preserved in ancient waste dumps and lava tubes). Around sixty of these species—nearly half the bird fauna—were honeycreepers, all descended from a single ancestral finch that arrived on the islands about four million years ago. Sadly, only twenty species of honey-creeper remain, all of them endangered. The rest were destroyed by hunting, habitat loss, and human-introduced predators like rats and mongooses. But even the few remaining honeycreepers show a fantastic diversity of ecological roles, as shown in figure 22. The bill of a bird can tell us a lot about its diet. Some species have curved bills for sipping nectar from flowers, others stout, parrotlike bills for cracking hard seeds or crushing twigs, still others thin pointy bills for picking insects from foliage, and some even have hooked bills for prying insects from trees, filling the role of a woodpecker. As on the Galápagos, we see one group that is overrepresented, with species filling niches occupied by very different species on continents or continental islands.
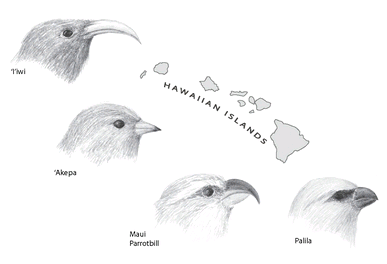
FIGURE 22
. An adaptive radiation: some related species of Hawaiian honeycreepers that evolved after their finchlike ancestor colonized the islands. Each finch has a bill that enables it to use different food. The ‘i’iwi’s slender bill helps it sip nectar from long tubular flowers, the ‘akepa has a slightly crossed bill that allows it to pry open buds to search for insects and spiders, the Maui parrotbill has a massive bill for prying up bark and splitting twigs to find beetle larvae, and the palila’s short but strong bill helps it open seed pods and extract the seeds.
. An adaptive radiation: some related species of Hawaiian honeycreepers that evolved after their finchlike ancestor colonized the islands. Each finch has a bill that enables it to use different food. The ‘i’iwi’s slender bill helps it sip nectar from long tubular flowers, the ‘akepa has a slightly crossed bill that allows it to pry open buds to search for insects and spiders, the Maui parrotbill has a massive bill for prying up bark and splitting twigs to find beetle larvae, and the palila’s short but strong bill helps it open seed pods and extract the seeds.
Oceanic islands also harbor radiations of plants and insects. St. Helena, though lacking many groups of insects, is home to dozens of species of small, flightless beetles, especially wood weevils. On Hawaii, the group that I study—fruit flies of the genus
Drosophila—
is positively luxuriant. Although the Hawaiian Islands make up only 0.004 percent of earth’s land, they contain nearly half of the world’s two thousand species of
Drosophila.
And then there are the remarkable radiations of plants in the sunflower family on the Juan Fernández archipelago and St. Helena, some of which have become small woody trees. Only on oceanic islands can small flowering plants, freed from competition with larger shrubs and trees, evolve into trees themselves.
Drosophila—
is positively luxuriant. Although the Hawaiian Islands make up only 0.004 percent of earth’s land, they contain nearly half of the world’s two thousand species of
Drosophila.
And then there are the remarkable radiations of plants in the sunflower family on the Juan Fernández archipelago and St. Helena, some of which have become small woody trees. Only on oceanic islands can small flowering plants, freed from competition with larger shrubs and trees, evolve into trees themselves.
So far we’ve learned two sets of facts about oceanic islands: they are missing many groups of species that live on continents and continental islands, and yet the groups that
are
found on oceanic islands are replete with many similar species. Together these observations show that, compared to other areas of the world, life on oceanic islands is
unbalanced.
Any theory of biogeography worth its salt has to explain this contrast.
are
found on oceanic islands are replete with many similar species. Together these observations show that, compared to other areas of the world, life on oceanic islands is
unbalanced.
Any theory of biogeography worth its salt has to explain this contrast.
But there’s something more here too. Take a look at the following list of the groups that are often native to oceanic islands and those that are usually missing (Juan Fernández is just one group of islands that conforms to the list):
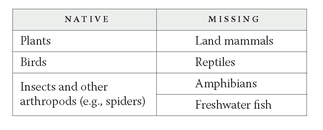
What’s the difference between the two columns? A moment’s thought gives the answer. Species in the first column can colonize an oceanic island through long-distance dispersal; species in the second column lack this ability. Birds are capable of flying great distances over the sea, carrying with them not only their own eggs but also seeds of plants they’ve eaten (which can germinate from their droppings), parasites in their feathers, and small organisms sticking to mud on their feet. Plants can get to islands as seeds, floating across expanses of sea. Seeds with barbs or sticky coverings can hitchhike to islands on the feathers of birds. The light spores of ferns, fungi, and mosses can be carried huge distances by the wind. Insects too can fly to islands or be taken by winds.
In contrast, animals in the second column have great difficulty crossing expanses of sea. Land mammals and reptiles are heavy and can’t swim very far. And most amphibians and freshwater fish simply can’t survive in salt water.
So the kinds of species that we find on oceanic islands are precisely those that can arrive across the sea from distant lands. But what is the evidence that they do so? Every ornithologist knows of occasional “visitor” birds found thousands of miles from their normal habitat, the victim of winds or faulty navigation. Some birds have even established breeding colonies on oceanic islands in historical times. The purple gallinule, long an occasional visitor to the remote island of Tristan da Cunha in the South Atlantic, finally started breeding there in the 1950s.
Darwin himself did some simple yet elegant experiments showing that seeds from some plant species could still germinate after prolonged immersion in seawater. Seeds from the West Indies have been found on the distant shores of Scotland, obviously carried by the Gulf Stream, and “drift seeds” from continents or other islands are also found on the shores of South Pacific islands. Caged birds can retain plant seeds in their digestive tracts for a week or more, showing the likelihood of long-distance transport. And there have been many successful attempts to sample insects in the air using traps attached to airplanes or ships far from land. Among the species collected have been locusts, moths, butterflies, flies, aphids, and beetles. Charles Lindbergh, on a 1933 trip across the Atlantic, exposed glass microscope slides to the air, capturing numerous microorganisms and insect parts. Many spiders disperse as juveniles by “ballooning” with parachutes of silk; these wanderers have been found several hundred miles from land.
Other books
Inspector of the Dead by David Morrell
Grendel Unit 2: Ignition Sequence by Bernard Schaffer
Koko Takes a Holiday by Kieran Shea
Ten Days by Janet Gilsdorf
The Secret Doctor by Joanna Neil
The Haunting (Immortals) by Robin T. Popp
Accidentally Demonic by Dakota Cassidy
The Doom of Kings: Legacy of Dhakaan - Book 1 by Don Bassingthwaite
Sins of the Highlander (A Highland Erotic Romance) by Halliday, Dawn
The Outlaw Josey Wales by Carter, Forrest