What Technology Wants (16 page)
Read What Technology Wants Online
Authors: Kevin Kelly

The notion of reproducibility in evolution is highly controversial. But since convergence is not only big news in biology but also strongly suggests convergence in the technium, it is worth looking at further evidence for it in nature. Depending on how one measures the concept of “independent,” the catalog of visible examples of independent, convergent evolution is hundreds long and counting. Any list will certainly include the three-time evolution of flapping wings in birds, bats, and pterodactyls (reptiles of the dinosaur era). The last common ancestor among these three lineages did not have wings, which means that each line evolved its wings independently. Despite their vast taxonomic distance, the wings in these three cases are remarkably similar in form: skin stretched over bony limbs. Navigation by echolocation has been found four times: in bats, dolphins, and two species of cave-dwelling birds (the South American oilbird and Asian swiftlet). Bipedality recurs in humans and birds. Antifreeze compounds were evolved twice in ice fish, once in the Artic and once in the Antarctic. Both hummingbirds and sphinx moths evolved to hover over flowers sucking nectar through a thin tube. Warm-bloodedness evolved more than once. Binocular vision evolved many times in distant taxa. Bryozoa, a family of coral, evolved distinctive helical colonies six different times over 400 million years. Social cooperation evolved in ants, bees, rodents, and mammals. Seven widely separated corners of the plant kingdom evolved insectivorous speciesâeating insects for nitrogen. Succulent leaves evolved multiple times across taxonomic distance, jet propulsion twice. Buoyant swim bladders evolved independently in many varieties of fish, mollusks, and jellyfish. Flapping wings constructed of taut membranes over skeleton frames arose more than once in the insect kingdom. While humans have technically evolved fixed-wing aircraft and spinning-wing aircraft, we haven't yet made a viable flapping-wing craft. On the other hand, fixed-wing gliders (flying squirrels, flying fish) and spinning-wing gliders (many seeds) have evolved a number of times. In fact, three species of rodentlike gliders also display convergence: the flying squirrel as well as the squirrel glider and marsupial sugar glider, both of Australia.
Because of its lone tectonic wanderings in geologic time, the continent of Australia is a laboratory for parallel evolution. There are multiple examples of marsupials in Australia paralleling placental mammals from the Old World, even in the past. Saber-canine teeth are found in both the extinct marsupial thylacosmilus and the extinct saber-toothed cat. Marsupial lions had retractable claws like feline cats.
Dinosaurs, our iconic distant cousins, independently evolved a number of innovations in parallel with our common vertebrate ancestors. In addition to the parallels between flying pterodactyls and bats, there were the streamlined ichthyosaurs that mirrored dolphins and mosasaurs, which paralleled whales. Triceratops evolved beaks similar to those of both parrots and octopus and squid. Snakelike pygopodidae were as legless as reptilian snakes later were.
The less taxonomic distance between lineages, the more commonâbut less significantâconvergence becomes. Both frogs and chameleons independently evolved rapid-fire “harpoon tongues” to snatch prey at a distance. All three major phyla of mushrooms have separately evolved species that produce dark, dense, underground, trufflelike fruits; and in North America alone there are more than 75 mushroom genera that include “truffles,” many of which evolved independently.
For some biologists, occurrences of convergence are merely a statistical curiosity, sort of like meeting someone else with your own name and birth date. Weird, but so what? Given enough species and enough time you are bound to encounter two that cross paths morphologically. But homologous features are actually the rule in biology. Most homology is invisible and occurs among related species. Relatives naturally share features, while the unrelated share fewer, so unrelated homology is more meaningful and noticeable. Either way, most methods used by life are used by more than one organism and in more than one phylum. What is rare is a trait that has
not
been reused somewhere in nature. Richard Dawkins challenged naturalist George McGavin to name biological “innovations” that have evolved only once, and McGavin was able to compile only a handful, such as the bombardier beetle, which mixes two chemicals on demand to shoot a noxious stream at enemies, or the diving-bell spider, which uses a bubble to breathe. Simultaneous, independent invention seems to be the rule in nature. As I argue in the next chapter, simultaneous, independent invention also seems to be the rule in the technium. In both realms, natural evolution and technological evolution, convergence creates inevitabilities. Inevitability is even more controversial than reproducibility and so demands yet more evidence.
not
been reused somewhere in nature. Richard Dawkins challenged naturalist George McGavin to name biological “innovations” that have evolved only once, and McGavin was able to compile only a handful, such as the bombardier beetle, which mixes two chemicals on demand to shoot a noxious stream at enemies, or the diving-bell spider, which uses a bubble to breathe. Simultaneous, independent invention seems to be the rule in nature. As I argue in the next chapter, simultaneous, independent invention also seems to be the rule in the technium. In both realms, natural evolution and technological evolution, convergence creates inevitabilities. Inevitability is even more controversial than reproducibility and so demands yet more evidence.
Return to the recurring eye. The retina is lined with a layer of a very specialized protein that performs the tricky work of perceiving light. This protein, called rhodopsin, transfers the photon energy from incoming light to an outgoing electrical signal sent along the optic nerve. Rhodopsin is an archaic molecule present not only in the retinas of camera eyes but also in the most primitive lensless eye spot of a lowly worm. It is found throughout the animal kingdom, and it retains its structure wherever it is found because it works so well. The same molecule has probably remained unchanged for billions of years. Several competing light-trigger molecules (cryptochromes) aren't as efficient or robust, suggesting that rhodopsin is simply the best molecule for seeing that can be found after two billion years of looking. But surprisingly, rhodopsin is another example of convergent evolution, because it evolved twice in two separate kingdoms in the deep pastâonce in Archaea and once in Eubacteria.
This fact should shock us. The number of possible proteins is astronomical. There is an alphabet of 20 base symbols (amino acids) that make up every protein “word,” which on average is, say, 100 symbols, or 100 bases, long. (In fact, many proteins are much longer, but for this calculation 100 is sufficient.) The total number of possible proteins that evolution could generate (or discover) is 100
20
or 10
39
. This means that there are more possible proteins than there are stars in the universe. But let's simplify that. Because only one in a million amino acid “words” folds into a functioning protein, let's vastly reduce that magnitude and agree that the number of potential working proteins is equal to the number of stars in the universe. Discovering a specific protein would be equivalent to randomly finding a specific star in the vastness of space.
20
or 10
39
. This means that there are more possible proteins than there are stars in the universe. But let's simplify that. Because only one in a million amino acid “words” folds into a functioning protein, let's vastly reduce that magnitude and agree that the number of potential working proteins is equal to the number of stars in the universe. Discovering a specific protein would be equivalent to randomly finding a specific star in the vastness of space.
By this analogy evolution finds new proteins (new stars) by a series of hops. It jumps from one protein to a “nearby” related one and then hops on to the next novel form until it reaches some remote unique protein far from where it started, just as one might travel to a distant sun by hopping stars. But in a universe as large as ours, once you landed on a distant star one hundred random hops away, you would never reach it again by the same random process. It is statistically impossible. But that is what evolution did with rhodopsin. Out of all the protein stars in the universe, it found this oneâa protein that has not been improved upon for billenniaâtwice.
And the impossibility of “twice-struck” keeps happening in life. Evolutionist George McGhee writes in a paper entitled “Convergent Evolution”: “The evolution of the ichthyosaur or porpoise morphology is not trivial. It can be correctly described as nothing less than astonishing that a group of land-dwelling tetrapods, complete with four legs and a tail, could devolve their appendages and their tails back into fins like those of a fish. Highly unlikely, if not impossible? Yet it happened twice, convergently in the reptiles and the mammals, two groups of animals that are not closely related. We have to go back in time as far as the Carboniferous to find a common ancestor for them; thus, their genetic legacies are very, very different. Nonetheless, the ichthyosaur and the porpoise both have independently re-evolved fins.”
What, then, guides this return to the improbable? If the same protein, or “contingent” form, is evolved twice, it is obvious that every step of the way cannot be random. The prime guidance for these parallel journeys is their common environment. Both archaea rhodopsin and eubacterial rhodopsin, and both ichthyosaur and dolphin, float in the same seas with the same advantages gained by adaptation. In the case of rhodopsin, because the molecular soup surrounding the precursor molecules is basically the same, the selection pressure will tend to favor the same direction on each hop. In fact, the match of environmental niche is usually the reason given for occurrences of convergent evolution. Arid, sandy deserts on different continents tend to produce largeeared, long-tailed, hopping rodents because the climate and terrain sculpts a similar set of pressures and advantages.
Yes, but why, then, doesn't every similar desert in the world produce a kangaroo rat, or jerboa, and why aren't all desert rodents some version of kangaroo rats? The orthodox answer is that evolution is a highly contingent process, where random events and pure luck change the course, so that even within parallel environments it is very rare to arrive at the same morphological solution. Contingency and luck are so strong in evolution that the marvel is that convergence ever happens. Based on the number of possible forms that can be assembled from the molecules of life and the central role of random mutation and deletion in shaping them, significant convergence from independent origins should be as scarce as miracles.
But a hundred, or a thousand, cases of isolated significant convergent evolution suggest something else at work. Some other force pushes the self-organization of evolution toward recurring solutions. A different dynamic besides the lottery of natural selection steers the course of evolution so that it can reach an unlikely remote destination more than once. It is not a supernatural force but a fundamental dynamic as simple at its core as evolution itself. And it is the same force that funnels convergence in technology and culture.
Evolution is driven toward certain recurring and inevitable forms by two pressures:
1. The negative constraints cast by the laws of geometry and physics, which limit the scope of life's possibilities.
2. The positive constraints produced by the self-organizing complexity of interlinked genes and metabolic pathways, which generate a few repeating new possibilities.
These two dynamics create a push in evolution that gives it a direction. Both of these two dynamics continue to operate in the technium as well and shape the inevitabilities along the course of the technium. Let me address each influence in turn, starting with the way chemistry and physics shape life and, by extension, the inventions of our mind in the technium.
Plants and animals come in a bewildering diversity of scales. Insects can be microscopic, like lice, or giant, like horned beetles the size of shoes; redwood trees tower 100 meters tall, and miniature alpine plants fit in a thimble; immense blue whales swell as big as ships, and pygmy chameleons shrink to less than an inch long. Yet the dimensions of each species are not arbitrary. They follow a scale ratio that is astonishingly constant in both plants and animals. This ratio is dictated by the physics of water. The strength of a cell wall is determined by the surface tension of water; that constant in turn mandates the maximum height per width of a body, any possible body. These physical forces play out not just on Earth, but everywhere in the universe, so we might expect any organisms based on water, whenever and wherever they evolve, to converge upon this same universal size ratio (adjusted for local gravity).
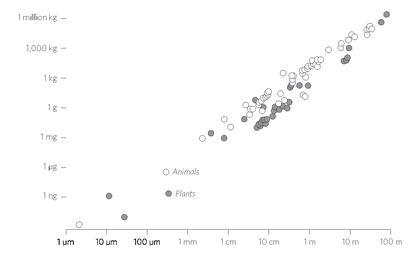
Size Ratio in Life.
The ratio between an organism's mass and its length is a constant in both plants and animals.
The ratio between an organism's mass and its length is a constant in both plants and animals.
The metabolism of life is likewise constrained. Small animals live fast and die young. Big animals plod along. The speed of life for animalsâthe rate at which their cells burn energy, the speed of their muscle twitches, the time it takes them to gestate or to matureâis remarkably proportional to their life span and size. Both metabolic rate and heart rate are proportional to the mass of the creature. These constants derive from the fundamental rules of physics and geometry and the natural advantages to minimizing energy surfaces (lung surface, cell surface, circulatory capacity, etc.). While a mouse's heart and lungs beat rapidly compared to an elephant's, both mouse and elephant count the same number of beats and breaths per life. It is as if mammals are assigned 1.5 billion heartbeats and told to use them as they like. Tiny mice speed ahead in a fast-forward version of an elephant's life.
In biology this constant ratio for metabolism was well known for mammals, but researchers recently realized a similar law governs all plants, bacteria, and even ecosystems. Dilute pools of cool oceanic algae can be thought of as a slow-motion version of a warm-blooded heart. The amount of energy per kilogram (or energy density) flowing through a plant or ecosystem is equivalent to metabolism. Many life processesâfrom the number of hours of sleep an animal needs to the hatching time for its eggs to the rate at which a forest accumulates wood mass to the mutation rate in DNAâall seem to follow a universal metabolism scaling law. “We've found that despite the incredible diversity of life, from a tomato plant to an amoeba to a salmon, once you correct for size and temperature, many of these [metabolic] rates and times are remarkably similar,” say James Gillooly and Geoffrey West, the researchers who discovered this law. “Metabolic rate is the fundamental biological rate,” they claimâ“a universal clock” reckoned in energy, the speed at which all life of any type proceeds. The clock is inevitable for anything living.
Other books
Loving A Romano by Lynn, Sindee
Elixir (Red Plague #1) (Red Plague Trilogy) by Abner, Anna
The Billionaire's Payment (BBW Erotic Romance) by Alexis Moore
A Lie Unraveled by Constance Masters
Southside (9781608090563) by Krikorian, Michael
The Law and Miss Penny by Sharon Ihle
Being Happy by David Tuffley
Smithy's Cupboard by Ray Clift
Aphrodite's Passion by Julie Kenner