The Act of Creation (67 page)
Read The Act of Creation Online
Authors: Arthur Koestler

developed' seems to be that it cannot be developed with the existing
theoretical tools without reducing the whole attempt to absurdity. It may
still be possible, and even respectable today for a geneticist to state
that: 'The hoary objection of the improbability of an eye or a hand or a
brain being evolved "by blind chance" has lost its force.' [5] But are
we also to assume that the behaviour-patterns of the digger-wasp, or of
the courtship and fighting rituals of various species have all evolved 'by
pure chance'? This assumption is implied in the doctrine of contemporary
genetics -- though rarely stated in explicit form. Similar assumptions
have been made by extreme behaviourists in the field of learning theory;
there is, in fact, a direct continuity between the doctrine of natural
selection operating on random mutations, and reinforcement operating
on random trials. Both grew out of the same philosophical climate. But
while learning theory is in full retreat from that extreme position,
and has a variety of alternative suggestions to offer, nothing the like
is in sight in the genetics of instinct-behaviour.
it is often difficult, if not impossible, to tell where the 'foundation'
ends and the 'building' starts. But the absence of fool-proof delineations
between 'inheritance', 'maturation', and 'learning' need not prevent us
from recognizing the existence of distinct patterns of animal behaviour
which are (a) stereotyped, (b) species-specific, (c) unlearnt in the
sense that they can be shown to appear, more or less completely, in
animals raised in isolation. It has been objected against this view that
'innate' behaviour, e.g. the pecking of chicks, may partly be due to
prenatal influences, [6] that 'isolation' is never absolute, [7] and
that learning may be practically instantaneous (as in imprinting). Such
arguments are valuable in showing that pure heredity sans environment is
an abstraction; but they do not alter the fact that each animal is born
with a hereditary potential to feed, hoard, court, nest, fight, and care
for its young in certain specific and highly characteristic ways which
are as much part of its native equipment as its morphological features,
and which can be modified by, but are not derived from, imitation and
learning. Only the unbalanced claims of some extreme behaviourists could
temporarily obscure the obvious fact that 'if the physical machinery for
behaviour develops under genetic control, then the behaviour it mediates
can scarcely be regarded as independent of inheritance'. [8]
experience. To quote the convergent definitions of one ethologist and
one psychologist: 'Learning is a central nervous process causing more
or less lasting changes in the innate behavioural mechanisms under
the influence of the outer world. . . .' [9] 'Learning is a process
by which an activity originates or is changed through reacting to
an encountered situation, provided that the characteristics of the
change in activity cannot be explained on the basis of native response
tendencies, maturation, or temporary states of the organism (fatigue,
drugs, etc.).' [10]
it is virtually impossible to draw a precise distinction between the
'innate' and 'acquired' aspects or components in the adult animal's
behaviour. Even the discrimination of biologically relevant sign-Gestalten
in the environment seems to require a minimum of experience; and
one must conclude, with Thorpe, that 'since comparison involves
learning, an element of learning enters into all orientation and all
perception. Accordingly it is suggested that the difference between inborn
and acquired behaviour is of degree rather than kind; it becomes, in fact,
a difference chiefly of degree of rigidity and plasticity.' [11] In
the terms of our schema, what is inherited is the specific and invariant
factor in the native skill -- its code. Its more or less flexible matrix
develops through learning from experience. To quote Thorpe again:
In each example of true instinctive behaviour there is a hard core
of absolutely fixed and relatively complex automatism -- an inborn
movement form. This restricted concept is the essence of the instinct
itself. Lorenz originally called it Erbkoordination or "fixed
action pattern". Such action patterns are items of behaviour in every
way as constant as anatomical structures, and are potentially just
as valuable for systematic, philogenetic studies. Every systematist
working with such groups as birds or higher insects will be able
to recall examples of the value of such fixed behaviour patterns in
classification. [12]
of fixed action-patterns, which incorporate the rules of the game of
courting, nest-building, duelling, etc. Each of these activities is again
a hierarchy of autonomous sub-skills. These tend to be more flexible
on the higher levels which co-ordinate the drive, and more rigid on the
lower levels. The autonomous sub-codes are restrained from spontaneous
activity by 'inhibitory blocks', and triggered into activity by patterned
impulses from higher echelons. This trigger-sensitive apparatus is called,
after Lorenz, the "innate releasing mechanism" -- or I.R.M. for short:
In all the channels which flow downward from the centre [of a given
drive], there is supposed to exist a physiological mechanism which
effectively prevents all discharge of activity unless the animal
encounters the right environmental situation and stimuli to remove
or release this block. Thus there is an innate releasing mechanism
(I.R.M.) . . . which is in some way attuned to the biologically right
stimulus in the environment . . . and which is, as it were, unlocked
by the appropriate releaser, thus allowing behaviour to proceed to the
next lower level. These in their turn incorporate blocks and, so long
as these remain, action of these lower centres cannot proceed.' [13]
is the reproductive behaviour of the male stickleback, which I shall
describe in the terminology of the present theory.
code', while hormonal activities provide the drive or motivation. The
fish then migrates into shallow water and swims around until a certain
environmental configuration (rise of temperature, combined with green
vegetation, etc.) strikes the 'right note', i.e. releases an efferent
impulse, which in turn triggers off the sub-code of the nest-building
activity. This activity is again subdivided into digging, glueing, etc.,
each of these skills governed by its autonomous sub-code. The latter are
activated by trigger releasers; the order of operations is determined by
inputs from the environment and proprioceptive feedbacks. The hierarchy of
mating behaviour remains blocked until nest-building is complete; but the
'fighting' hierarchy (with its five different sub-codes) may be called
at any moment into action by a trigger mechanism sensitive to a specific
sign-Gestalt input: 'red male entering territory'. In this case the
fighting code dominates the animal's entire behaviour, and nest-building
as well as other activities are inhibited while the emergency lasts:
the (functional) part monopolizes the attention of the whole.
The sub-units of the behaviour-pattern tend to become more specific
regarding input and more stereotyped in output on the lower levels of
the hierarchy. 'Which one of the five motor responses belonging to the
fighting pattern will be shown depends on sign stimuli that are still more
restricted in effect.' [14] The nuptial colours of the fish are shining
blue eyes and a red underbelly. Accordingly, any crude model which is
red underneath will release an attack, regardless of shape and size --
whereas a perfectly shaped model without nuptial colouring will not
do so. Apart from colour, behaviour also acts as a releaser. 'When the
stranger bites, the owner of the territory will bite in return. When the
stranger threatens, the owner will threaten back; when the stranger flees,
the owner will chase it; and so on. . . . [15] But fighting is rarer than
threat. The threat-behaviour of male sticklebacks is peculiar. Not only do
they dart towards the opponent with raised dorsal spines and open mouth,
ready to bite, but, when the opponent does not flee at once but resists,
the owner of the territory does not actually bite but points its head
down and, standing vertically in the water, makes some jerky movements
as if it were going to bore its snout into the sand. [16]
This of course is an exceptional example -- nest-building is a rarity
among fish. But the rigidity of fixed action patterns in certain
classes -- such as birds and insects -- remains nevertheless a striking
phenomenon. The ritualized rules of the game of courtship and display,
of threat and danger signals, of tournament fighting and social behaviour,
sometimes reminds one of the ceremonious observances at Byzantine courts,
at other times of the obsessive rituals of compulsion-neurotics. And
the process of 'ritualization' does indeed suggest the 'emancipation'
-- or isolation -- of a behaviour-pattern from its original context,
accompanied by intensification, stabilization, and rhythmic repetitiveness
of the pattern; the reasons are as yet hardly understood. [17]
Appetitive Behaviour and Consummatory Act
In spite of the relatively stereotyped nature of fixed action-patterns --
of which ritualization is an extreme example -- it would be entirely
wrong to regard the hierarchy of instinct behaviour as a one-way
affair, in which a plastic, general drive (the 'appetitive behaviour')
discharges downward along pre-formed and discrete alternative channels
into the completely rigid and mechanical, fixed-action-patterns of the
'consummatory act'. This conception of the organism as an automaton whose
'adaptability' is reduced to that of a kind of automatic record-changer
or jukebox, with a choice between a few dozen fixed records appropriate
to the occasion, seems to have originated in a misunderstanding of the
distinction made by Wallace Craig between 'appetitive behaviour' and
'consummatory act'. This point must be briefly discussed as it is of
some importance for the sections which follow.
Appetite (or 'appetance') was defined by Craig (1918) as a 'state of
agitation', a striving for an absent 'appeted' stimulus (conversely,
a striving to escape from a noxious or disturbing stimulus); whereas the
'cousummatory act' was meant to bring the activity to a close by attaining
(or escaping from) the appetitive stimulus, 'after which the appetitive
behaviour ceases and is succeeded by a state of relative rest'. [18]
More generally, 'the term appetitive behaviour is used by present-day
writers on ethology to mean the flexible or variable introductory phase
of an instinctive behaviour pattern or sequence'. [19]
Thus 'appetitive behaviour' became a more refined and noncommittal name
for the old, shop-soiled concepts of 'need', 'drive', 'instinct', and
'purpose'.* So far all was well; it was the 'consummatory act', which
led instinct-theory into a cul-de-sac. The trouble started, rather
inconspicuously, when first Woodworth [20] then, independently from
each other, K. S. Lashley and Konrad Lorenz became impressed with the
stereotyped character of certain 'consummatory acts' (animal rituals
and automatized habits in humans), as compared with the more general
'appetitive behaviour' or drive.** Eventually the focussing of attention
on such fixed patterns of behaviour led to a distortion of the whole
picture: Lorenz and Tinbergen made a rigid distinction between appetitive
behaviour which was supposed to be flexible, and consummatory acts which
were supposed to be completely fixed and automatic. Thus Tinbergen:
It will be clear, therefore, that this distinction between appetitiveBut what exactly, one might ask, constitutes a 'consummatory act'?
behaviour and consummatory act separates the behaviour as a whole into
two components of entirely different character. The consummatory
act is relatively simple; at its most complex it is a chain of
reactions. . . . But appetitive behaviour is a true purposive activity,
offering all the problems of plasticity, adaptiveness, and of complex
integration that baffle the scientist in his study of behaviour as
a whole. . . . Lorenz has pointed out . . . that purposiveness, the
striving towards an end, is typical only of appetitive behaviour and
not of consummatory actions. . . . Whereas the consummatory act seems
to be dependent on the centres of the lowest level of instinctive
behaviour, appetitive behaviour may be activated by centres of all
the levels above that of the consummatory act. . . . [21] The centres
of the higher levels do control purposive behaviour which is adaptive
with regard to the mechanisms it employs to attain the end. The lower
levels, however, give rise to increasingly simple and more stereotyped
movements, until at the level of the consummatory act we have to do
with an entirely rigid component, the taxis, the variability of which,
however, is entirely dependent on changes in the outer world. This
seems to settle the controversy; the consummatory act is rigid, the
higher patterns are purposive and adaptive. [22]
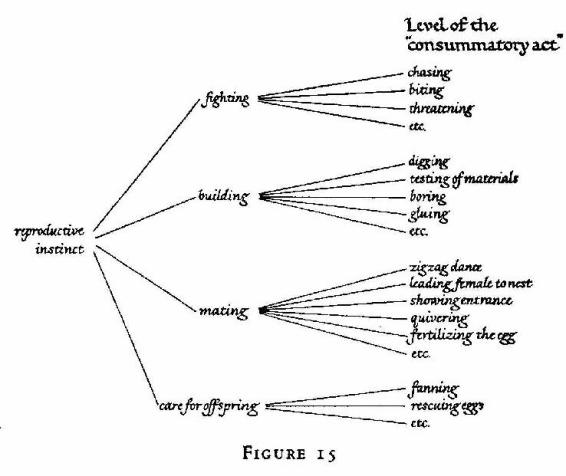
A glance at Tinbergen's diagram on
Other books
Katy's Men by Carr, Irene
02 Thunder of Heaven: A Joshua Jordan Novel by Tim Lahaye
Khyber Run by Amber Green
The Dark Clue by James Wilson
Latimer's Law by Mel Sterling
Creatus (Creatus Series) by Carmen DeSousa
Winning Her Over by Alexa Rowan
Silt, Denver Cereal Volume 8 by Claudia Hall Christian
The Troubled Air by Irwin Shaw
Tales of the Djinn: The Double by Emma Holly