Hen’s Teeth and Horse’s Toes (33 page)
Read Hen’s Teeth and Horse’s Toes Online
Authors: Stephen Jay Gould

If a unity of basic architecture underlies the diversity of zebra striping, then we must suspect that we are confronting a general pattern in nature, not just the “supreme abnormality” that Marianne Moore described. Darwin viewed horses in this light and recognized that the capacity for striping in all horses constituted a powerful argument for evolution itself. If zebras are odd and perfect adaptations for camouflage, God might have made them as we find them. But if zebras merely actuate and exaggerate a potential property of all horses, then the occasional realization of striping in other horses—where it cannot be viewed as a perfected adaptation ordained by God—must indicate a community of evolutionary descent.
Darwin devoted much space in chapter 5 of the
Origin of Species
to an exhaustive tabulation of occasional striping in other horses. Asses, he found, often have “very distinct transverse bars…like those on the legs of a zebra.” True horses often possess a spinal stripe, and some also have transverse leg bars. Darwin found a Welsh pony with three parallel stripes on each shoulder. And he noted that hybrids (with no zebra parents) were often strongly striped—an example of the common, and still mysterious, observation that hybrids often display ancestral reminiscences present in neither parent. “I once saw a mule,” Darwin wrote, “with its legs so much striped that any one at first would have thought that it must have been the product of a zebra.”
From this illustration of common, and often nonadaptive, patterns in all horses, Darwin drew one of his most powerful and passionate arguments for evolution—well worth quoting
in extenso:
He who believes that each equine species was independently created, will, I presume, assert that each species has been created with a tendency to vary, both under nature and under domestication, in this particular manner, so as often to become striped like other species of the genus; and that each has been created with a strong tendency, when crossed with species inhabiting distant quarters of the world, to produce hybrids resembling in their stripes, not their own parents, but other species of the genus. To admit this view is, as it seems to me, to reject a real for an unreal, or at least for an unknown, cause. It makes the works of God a mere mockery and deception; I would almost as soon believe with the old and ignorant cosmogonists, that fossil shells had never lived, but had been created in stone so as to mock the shells now living on the seashore.
The same theme also suggests an answer to the title of essay 28: “What, If Anything, Is a Zebra?” I advanced the argument that zebras may not form a group of closest relatives but a set of different horses that had either evolved stripes independently or inherited them from a common ancestor (while asses and true horses lost them). Bard’s hypothesis lends support to this conjecture because it suggests that the underlying pattern of zebra striping may be so simple that all horses include it in their repertoire of development. Zebras, then, may be the realization of a potential possessed by all horses.
Finally, moving from the sublime to the merely interesting, Bard proposes a solution to the primal dilemma and argues that zebras are black animals with white stripes after all. The white underbelly, he points out, is a lousy argument because many fully colored mammals are white underneath. Color may be generally inhibited in this region for reasons at present unknown. Mammals do not have their colors painted on a white background. The basic issue may then be rephrased: Does striping result from an inhibition or a deposition of melanin? If the first, zebras are black animals; if the second, they are white with black stripes.
Biologists often look to teratologies, or abnormalities of development, to solve such issues. Bard has uncovered an abnormal zebra whose “stripes” are rows of dots and discontinuous blotches, rather than coherent lines of color. The dots and blotches are white on a black background. Bard writes: “It is only possible to understand this pattern if the white stripes had failed to form properly and that therefore the ‘default’ color is black. The role of the striping mechanism is thus to inhibit natural pigment formation rather than to stimulate it.” The zebra, in other words, is a black animal with white stripes.
AS DARWIN CATALOGED
cases of striped horses and asses to illustrate a common ancestry with zebras (see last essay), he inevitably encountered one of the most famous animals of nineteenth-century natural history—the Earl of Morton’s mare. Darwin wrote in the
Origin of Species:
“In Lord Moreton’s [
sic
] famous hybrid from a chestnut mare and male quagga, the hybrid, and even the pure offspring subsequently produced from the mare by a black Arabian sire, were much more plainly barred across the legs than is even the pure quagga.”
The quagga, a zebra with stripes restricted to its neck and forequarters, is now extinct. It was not thriving in the early nineteenth century either, when the good Earl hoped to save the species by domesticating it. He was able to procure a male for his noble experiment but could never obtain a female. So he bred his male quagga with “a young chestnut mare of seven-eighths Arabian blood,” and obtained a hybrid with “very decided indications of her mixed origin.” Nothing surprising so far.
But the disappointed Lord Morton, unable to find more quaggas, sold his Arabian mare to Sir Gore Ouseley, who proceeded to breed her with “a very fine black Arabian horse.” When Lord Morton visited his friend and viewed the two offspring of pedigreed Arabian parents, he was astonished to note in them what he took to be a “striking resemblance to the quagga.” Somehow, the quagga father had influenced subsequent offspring sired by other males years after his permanent departure from the life of Lord Morton’s mare. How could such an influence be maintained long after physical contact had ceased?
Lord Morton’s mare was the most celebrated, but by no means the only, case of a phenomenon that German biologist August Weismann later named “telegony,” from Greek roots meaning “offspring at a distance,” or the idea that sires could influence subsequent progeny not fathered by them. Since the supposed phenomenon turned out to be an illusion, telegony now reposes as one more forgotten item on history’s ash heap, and neither Lord Morton nor his mare retain any notoriety today.
But historian of science Richard W. Burkhardt, Jr., who recently wrote an excellent article on the history of telegony in general and Lord Morton’s mare in particular (see bibliography), has demonstrated that telegony was once a major subject for research and “inspired the most extensive work in experimental animal breeding conducted in Britain between Darwin’s death in 1882 and the rediscovery of Mendel’s law in 1900.” Darwin himself was a major supporter of telegony.
If the supposed causes of telegony are a bit mysterious, Darwin’s reasons for espousing the idea may seem equally difficult to fathom. After all, he first discussed the progeny of Lord Morton’s mare in a context that implied an explanation opposed to telegony. As I wrote in the first two parts of this trilogy, Darwin had cataloged all cases he could find of asses and true horses with stripes. He used these striped horses as an effective argument for evolution: If God had created true horses, asses, and zebras as separate forms, why should we find occasional striped individuals in species that normally lack them? Does not this latent tendency for striping in all horses (permanently actualized only in zebras) indicate common descent? Why, then, did Darwin later implicate the previous quagga sire in the striping of subsequent offspring from Lord Morton’s mare? In this first discussion, from the
Origin of Species
, Darwin set out to prove that true horses and asses can develop stripes
without
any zebra influence. As we shall see, this original explanation was apparently correct.
Burkhardt argues that Darwin changed his mind and supported telegony because it fit so well with the (ironically) “non-Darwinian” theory of heredity that he developed in 1868. Under this “provisional hypothesis of pangenesis” (as Darwin called it), all cells of the body produce tiny particles, called gemmules, that course throughout the body, gather in the sex cells, and eventually transmit the characters of parents to offspring. Since gemmules might be altered if the cells producing them are changed by the influence of environment or the activity of animals themselves, acquired characters can be inherited and evolution has an important Lamarckian aspect.
Telegony meshed well with pangenesis because gemmules included with the quagga’s sperm would have remained in the body of Lord Morton’s mare and extended their influence to her subsequent offspring. (Darwin even once speculated that gemmules passed in sperm might explain why some women grow to resemble their husbands. As to why so many people resemble their dogs, Darwin maintained a discreet silence.)
Telegony finally fell when a new theory of heredity rose to favor and excluded it. August Weismann, who championed the strict Darwinism of natural selection against all forms of Lamarckian inheritance (including Darwin’s own pangenesis), argued for what he called “the continuity of germ plasm.” He held that reproductive cells are completely isolated from the rest of the body and cannot be influenced by whatever forces mold and alter other organs and tissues. Acquired characters cannot affect the next generation because they cannot penetrate the “casket” that holds reproductive cells and transmits itself in toto from generation to generation. (The fertilized egg, of course, is formed by the union of two reproductive cells. As it starts to divide, however, the nonreproductive cells form, eventually develop into the organism’s body, and become rigidly segregated from the continuous lineage of germ cells. Telegony makes no sense because, even if they existed, male gemmules in a female’s body could not get to the germ cells—unless they managed to reach the ovaries themselves and modify the immature ova.)
Telegony kicked about in scientific literature for seventy years, from Morton’s note to the Royal Society in 1820 until Weismann’s challenge. When Weismann proposed the continuity of germ plasm, telegony became a threatening anomaly demanding affirmation or rejection. Many tests were made, and telegony failed them miserably. In particular, J. C. Ewart, Regius Professor of Natural History at Edinburgh, tried to repeat Morton’s own experiment. Since quaggas had joined
Eohippus
in the realm of departed horses, Ewart mated twenty mares of different races and breeds with a male Burchell’s zebra. The first hybrid, born in 1896, had stripes as expected. Ewart then mated the mare with a second sire, an Arab stallion. Their offspring also had stripes, albeit faint, and telegony seemed vindicated. But Ewart knew that he needed controls and therefore bred the same Arab stallion to other mares that “had never so much as seen a zebra.” The offspring of these matings were as richly striped as the foal from the mare that had previously mated with a zebra. Darwin had been right the first time. Stripes do not arise from a mysterious previous zebra influence; they represent a potential pathway of development for all horses.
I have not recounted this tale of telegony for its own sake, since antiquarian musings only excite professionals and trivia buffs. Rather, as Burkhardt emphasizes, the story embodies a larger, troubling, and important issue about the nature of fact in science. Telegony, so far as we can tell, was wrong; yet it remained in the literature as a pristine fact, largely unchallenged, for seventy years. In a reversal of the stereotyped scenario, where a single sturdy fact arises to destroy an entire edifice of theory, the “fact” of telegony came first, became entrenched, and was only seriously challenged when a theory—Weismann’s continuity of germ plasm—rendered it anomalous. Burkhardt notes that in the usual model “a long-accepted theory is toppled by a newly discovered, apparently anomalous fact. In the case of telegony, in contrast, a long-accepted ‘fact’ was discredited when confronted with a new, apparently contradictory theory.”
In part, Burkhardt suggests, telegony gathered favor because it fit well with a variety of nineteenth-century assumptions, ranging from the “natural” dominance of males over females to support for separation of races by the argument that sexual contact with a lower race might extend its baleful influence far beyond the immediate consequences of the act itself. In part, telegony simply wasn’t controversial enough and nobody bothered to test Lord Morton’s improbable assertion. When tested, telegony fell, and to this extent the usual model of science as objective experiment was vindicated. But another aspect of the conventional view did not apply: fact did not act as the cleansing broom for outmoded theory. Rather, a false “fact” endured for an uncomfortably long time until theory demanded its test. What does this story tell us about the relation of fact and theory in science and about the role of single, isolated facts in the first place? I will return to these generalities, but first, another and similar tale.
In 1922, British paleontologist A. E. Trueman published the most famous paper of our century on a supposedly unbroken lineage of evolving fossils. He argued that flat oysters had slowly evolved into coiled oysters of the genus
Gryphaea
. Although coiling was originally advantageous in raising the oysters above an increasingly muddy sea floor, the trend, once started, could not be halted.
Gryphaea
built one coiled valve and one flat valve lying like a cap atop its coiling partner. Eventually, the coil grew up and over the cap, finally pressing hard upon it. Unable to open its shell,
Gryphaea
perished, imprisoned in its own embrace.
When Trueman published his paper, most paleontologists were not Darwinians. The theory of orthogenesis, or “straight line” evolution forcing organisms along predetermined paths, was still popular. An inexorable trend that natural selection could not stop and that led to the demise of a lineage was an expected phenomenon. Thus, Trueman’s story was not challenged by paleontologists, and like the zebra’s impact on Lord Morton’s mare, overcoiled
Gry
-
phaea
became an established fact.
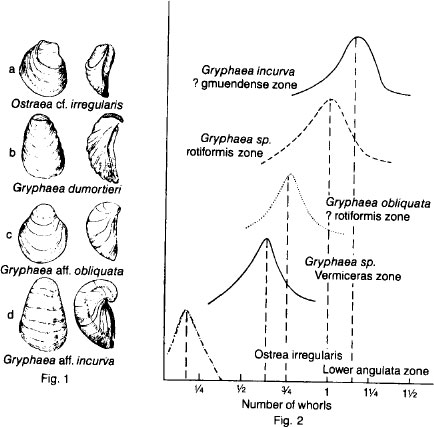
Supposed increase of coiling in
Gryphaea
according to Trueman. Ancestors (top) to coiled descendants (bottom) in pictures of oysters to left. Ancestors (bottom) to coiled descendants (top) in charts of variation in number of coils to right.
FROM K. JOYSEY, BIOLOGICAL REVIEWS, VOL
., 34, 1959.
In the late 1930s, after a gestation of nearly one hundred years from Darwin’s great insight and eighty years from its publication, natural selection finally triumphed as the accepted theory of evolutionary change, and
Gryphaea
became an anomaly. How could a lineage
actively
evolve itself to extinction if evolutionary change is directed by natural selection and can, therefore, only occur in directions that adapt organisms to local environments? (Extinction based upon an inability to change rapidly enough in the face of environmental perturbation is another matter and perfectly orthodox in a Darwinian world.)
Darwinians reacted to
Gryphaea
in a variety of ways, but all were deeply troubled. Some, like J. B. S. Haldane, simply admitted puzzlement: “The exaggerated coiling of
Gryphaea
cannot at present be explained with any strong degree of likelihood.” Others, like G. G. Simpson, tried to steer around the problem with faintly plausible, but evidently ad hoc suggestions: since overcoiling only affected the oldest, and probably postreproductive, individuals, it may even have benefited the population by clearing out the old codgers and making space for the young and vigorous. Yet, through all these efforts in salvaging natural selection, no one asked the more basic question: Is it even true in the first place? Overcoiling in
Gryphaea
had become a fact.
But it is not true; overcoiling in
Gryphaea
is as chimerical as the supposed influence of Lord Morton’s quagga upon the subsequent progeny of his mare. In 1959, Anthony Hallam, now a good friend and professor of geology at Birmingham, but then a brash graduate student, wrote an iconoclastic paper with two controversial claims: first, that
Gryphaea
had not evolved from flat oysters at all, but had migrated into southern England from elsewhere; and second, that
Gryphaea
arrived in England as coiled as it would ever be—and that no trend to increased coiling within
Gryphaea
could be demonstrated at all. Hallam’s conclusion scandalized many senior scientists who had known Trueman. H. H. Swinnerton, dean of British paleontology, wrote back in anger, accusing Hallam of both “sin” and “monstrous error,” an odd pair of accusations for a scientific paper.
Twenty years later the dust has settled, and I do not think that anyone would now doubt Hallam’s debunking. For Hallam’s first claim—that
Gryphaea
did not evolve from flat oysters—H. B. Stenzel’s impressive monograph (
Treatise on Invertebrate Paleontology
, part N, 1971) has proved that the lineages of flat oysters (genus
Ostrea
) and coiled oysters (genus
Gryphaea
) have been separate from their first appearance, that
Gryphaea
is a bit older than
Ostrea
, and that coiled ancestors for Trueman’s
Gryphaea
were living in Greenland before
Ostrea
ever appeared in England.